正在加载图片...
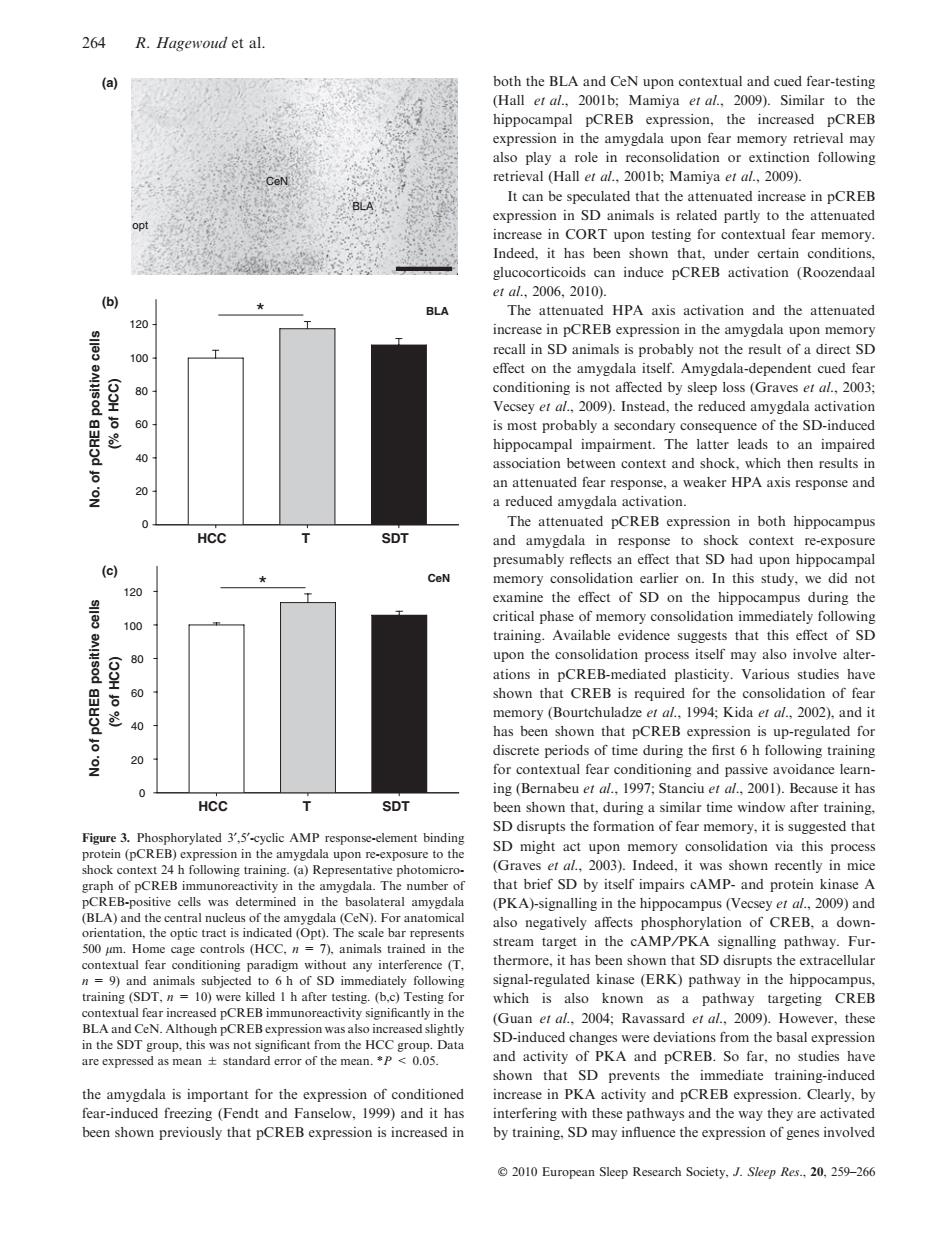
264 R.Hagewoud et al. (a) both the BLA and CeN upon contextual and cued fear-testing (Hall et al.,2001b;Mamiya et al.,2009).Similar to the hippocampal pCREB expression,the increased pCREB expression in the amygdala upon fear memory retrieval may also play a role in reconsolidation or extinction following retrieval (Hall et al.,2001b:Mamiya et al.,2009) It can be speculated that the attenuated increase in pCREB expression in SD animals is related partly to the attenuated 001 increase in CORT upon testing for contextual fear memory. Indeed.it has been shown that,under certain conditions. glucocorticoids can induce pCREB activation (Roozendaal e1al.2006.2010). (b) BLA The attenuated HPA axis activation and the attenuated 120 increase in pCREB expression in the amygdala upon memory recall in SD animals is probably not the result of a direct SD 100 effect on the amygdala itself.Amygdala-dependent cued fear 80 conditioning is not affected by sleep loss (Graves et al.,2003: Vecsey et al.,2009).Instead,the reduced amygdala activation 60 is most probably a secondary consequence of the SD-induced hippocampal impairment.The latter leads to an impaired 40 association between context and shock,which then results in 20 an attenuated fear response,a weaker HPA axis response and a reduced amygdala activation. The attenuated pCREB expression in both hippocampus HCC SDT and amygdala in response to shock context re-exposure presumably reflects an effect that SD had upon hippocampal (c) ★ CeN memory consolidation earlier on.In this study,we did not 120 examine the effect of SD on the hippocampus during the critical phase of memory consolidation immediately following 100 training.Available evidence suggests that this effect of SD 80 upon the consolidation process itself may also involve alter- (53HJ ations in pCREB-mediated plasticity.Various studies have 60 shown that CREB is required for the consolidation of fear memory (Bourtchuladze et al.,1994;Kida er al.,2002).and it has been shown that pCREB expression is up-regulated for discrete periods of time during the first 6 h following training 20 for contextual fear conditioning and passive avoidance learn- 0 ing(Bernabeu et al.,1997;Stanciu et al.,2001).Because it has HCC SDT been shown that.during a similar time window after training. SD disrupts the formation of fear memory,it is suggested that Figure 3.Phosphorylated 3,5'-cyclic AMP response-element binding protein(pCREB)expression in the amygdala upon re-exposure to the SD might act upon memory consolidation via this process shock context 24 h following training.(a)Representative photomicro- (Graves et al.,2003).Indeed,it was shown recently in mice graph of pCREB immunoreactivity in the amygdala.The number of that brief SD by itself impairs cAMP-and protein kinase A pCREB-positive cells was determined in the basolateral amygdala (PKA)-signalling in the hippocampus(Vecsey et al.,2009)and (BLA)and the central nucleus of the amygdala(CeN).For anatomical also negatively affects phosphorylation of CREB,a down- orientation,the optic tract is indicated (Opt).The scale bar represents 500 pm.Home cage controls (HCC.n 7).animals trained in the stream target in the cAMP/PKA signalling pathway.Fur- contextual fear conditioning paradigm without any interference (T, thermore,it has been shown that SD disrupts the extracellular n=9)and animals subjected to 6 h of SD immediately following signal-regulated kinase (ERK)pathway in the hippocampus, training (SDT,n 10)were killed I h after testing.(b,c)Testing for which is also known as a pathway targeting CREB contextual fear increased pCREB immunoreactivity significantly in the (Guan et al..2004:Ravassard et al..2009).However,these BLA and CeN.Although pCREBexpression was also increased slightly in the SDT group,this was not significant from the HCC group.Data SD-induced changes were deviations from the basal expression are expressed as mean+standard error of the mean.*P 0.05. and activity of PKA and pCREB.So far,no studies have shown that SD prevents the immediate training-induced the amygdala is important for the expression of conditioned increase in PKA activity and pCREB expression.Clearly,by fear-induced freezing(Fendt and Fanselow,1999)and it has interfering with these pathways and the way they are activated been shown previously that pCREB expression is increased in by training,SD may influence the expression of genes involved 2010 European Sleep Research Society,J.Sleep Res..20,259-266the amygdala is important for the expression of conditioned fear-induced freezing (Fendt and Fanselow, 1999) and it has been shown previously that pCREB expression is increased in both the BLA and CeN upon contextual and cued fear-testing (Hall et al., 2001b; Mamiya et al., 2009). Similar to the hippocampal pCREB expression, the increased pCREB expression in the amygdala upon fear memory retrieval may also play a role in reconsolidation or extinction following retrieval (Hall et al., 2001b; Mamiya et al., 2009). It can be speculated that the attenuated increase in pCREB expression in SD animals is related partly to the attenuated increase in CORT upon testing for contextual fear memory. Indeed, it has been shown that, under certain conditions, glucocorticoids can induce pCREB activation (Roozendaal et al., 2006, 2010). The attenuated HPA axis activation and the attenuated increase in pCREB expression in the amygdala upon memory recall in SD animals is probably not the result of a direct SD effect on the amygdala itself. Amygdala-dependent cued fear conditioning is not affected by sleep loss (Graves et al., 2003; Vecsey et al., 2009). Instead, the reduced amygdala activation is most probably a secondary consequence of the SD-induced hippocampal impairment. The latter leads to an impaired association between context and shock, which then results in an attenuated fear response, a weaker HPA axis response and a reduced amygdala activation. The attenuated pCREB expression in both hippocampus and amygdala in response to shock context re-exposure presumably reflects an effect that SD had upon hippocampal memory consolidation earlier on. In this study, we did not examine the effect of SD on the hippocampus during the critical phase of memory consolidation immediately following training. Available evidence suggests that this effect of SD upon the consolidation process itself may also involve alterations in pCREB-mediated plasticity. Various studies have shown that CREB is required for the consolidation of fear memory (Bourtchuladze et al., 1994; Kida et al., 2002), and it has been shown that pCREB expression is up-regulated for discrete periods of time during the first 6 h following training for contextual fear conditioning and passive avoidance learning (Bernabeu et al., 1997; Stanciu et al., 2001). Because it has been shown that, during a similar time window after training, SD disrupts the formation of fear memory, it is suggested that SD might act upon memory consolidation via this process (Graves et al., 2003). Indeed, it was shown recently in mice that brief SD by itself impairs cAMP- and protein kinase A (PKA)-signalling in the hippocampus (Vecsey et al., 2009) and also negatively affects phosphorylation of CREB, a downstream target in the cAMP ⁄ PKA signalling pathway. Furthermore, it has been shown that SD disrupts the extracellular signal-regulated kinase (ERK) pathway in the hippocampus, which is also known as a pathway targeting CREB (Guan et al., 2004; Ravassard et al., 2009). However, these SD-induced changes were deviations from the basal expression and activity of PKA and pCREB. So far, no studies have shown that SD prevents the immediate training-induced increase in PKA activity and pCREB expression. Clearly, by interfering with these pathways and the way they are activated by training, SD may influence the expression of genes involved opt CeN BLA CeN HCC T SDT 0 20 40 60 80 100 120 * HCC T SDT No. of pCREB positive cells (% of HCC) No. of pCREB positive cells (% of HCC) 0 20 40 60 80 100 120 * BLA (a) (c) (b) Figure 3. Phosphorylated 3¢,5¢-cyclic AMP response-element binding protein (pCREB) expression in the amygdala upon re-exposure to the shock context 24 h following training. (a) Representative photomicrograph of pCREB immunoreactivity in the amygdala. The number of pCREB-positive cells was determined in the basolateral amygdala (BLA) and the central nucleus of the amygdala (CeN). For anatomical orientation, the optic tract is indicated (Opt). The scale bar represents 500 lm. Home cage controls (HCC, n = 7), animals trained in the contextual fear conditioning paradigm without any interference (T, n = 9) and animals subjected to 6 h of SD immediately following training (SDT, n = 10) were killed 1 h after testing. (b,c) Testing for contextual fear increased pCREB immunoreactivity significantly in the BLA and CeN. Although pCREB expression was also increased slightly in the SDT group, this was not significant from the HCC group. Data are expressed as mean ± standard error of the mean. *P < 0.05. 264 R. Hagewoud et al. 2010 European Sleep Research Society, J. Sleep Res., 20, 259–266�