正在加载图片...
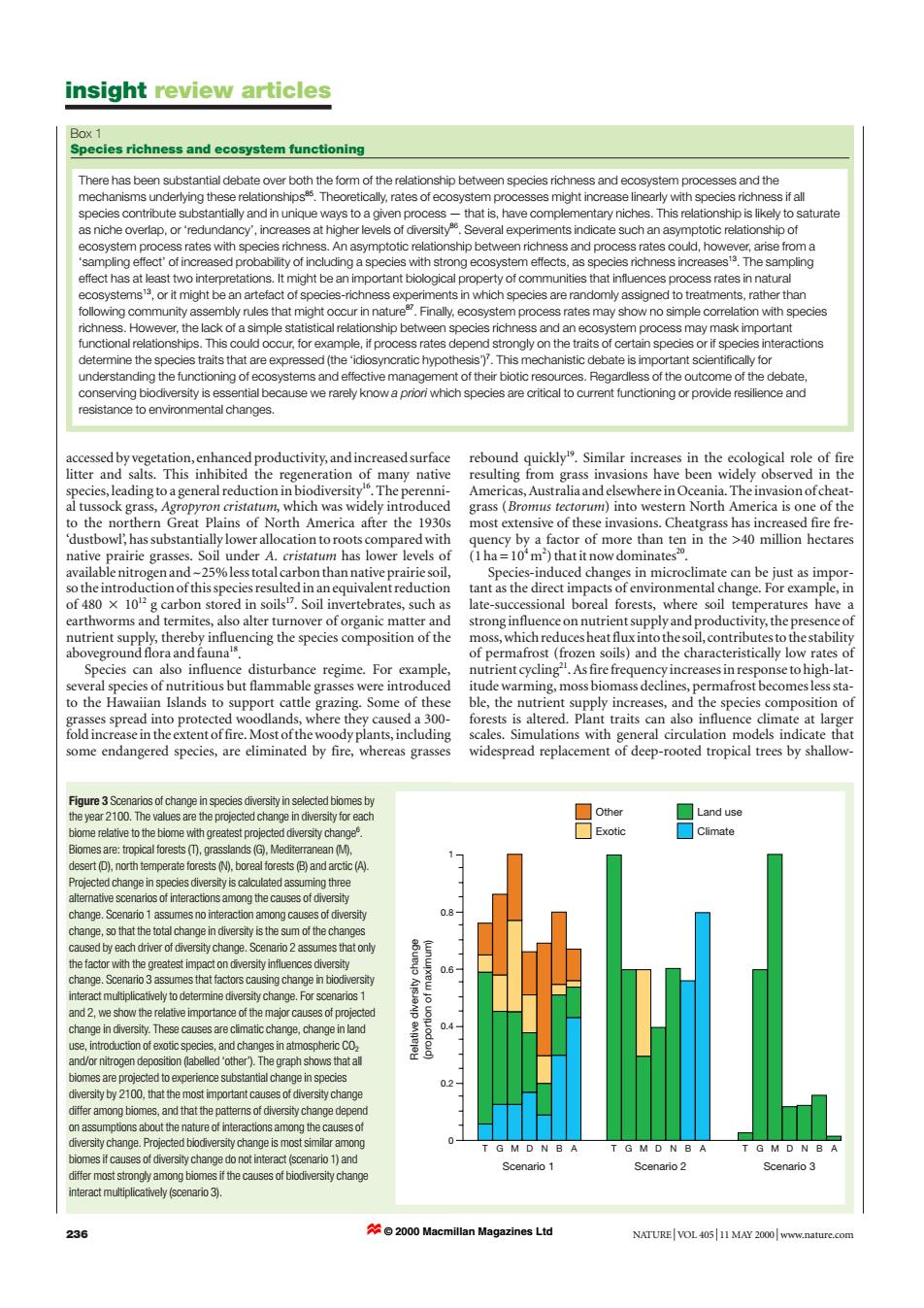
insight review articles Box 1 Species richness and ecosystem functioning There has been substantial debate over both the form of the relationship between species richness and ecosystem processes and the mechanisms underlying these relationships5.Theoretically,rates of ecosystem processes might increase linearly with species richness if all species contribute substantially and in unique ways to a given process-that is,have complementary niches.This relationship is likely to saturate as niche overlap,or'redundancy',increases at higher levels of diversity.Several experiments indicate such an asymptotic relationship of ecosystem process rates with species richness.An asymptotic relationship between richness and process rates could,however,arise from a 'sampling effectof increased probability of including a species with strong ecosystem effects,as species richness increasest.The sampling effect has at least two interpretations.It might be an important biological property of communities that influences process rates in natural ecosystems13,or it might be an artefact of species-richness experiments in which species are randomly assigned to treatments,rather than following community assembly rules that might occur in nature.Finally,ecosystem process rates may show no simple correlation with species richness.However,the lack of a simple statistical relationship between species richness and an ecosystem process may mask important functional relationships.This could occur,for example,if process rates depend strongly on the traits of certain species or if species interactions determine the species traits that are expressed (the'idiosyncratic hypothesis').This mechanistic debate is important scientifically for understanding the functioning of ecosystems and effective management of their biotic resources.Regardless of the outcome of the debate, conserving biodiversity is essential because we rarely know a priori which species are critical to current functioning or provide resilience and resistance to environmental changes. accessed by vegetation,enhanced productivity,and increased surface rebound quickly.Similar increases in the ecological role of fire litter and salts.This inhibited the regeneration of many native resulting from grass invasions have been widely observed in the species,leading to ageneral reduction in biodiversity.The perenni- Americas,Australia and elsewhere in Oceania.The invasion ofcheat- al tussock grass,Agropyron cristatum,which was widely introduced grass(Bromus tectorum)into western North America is one of the to the northern Great Plains of North America after the 1930s most extensive of these invasions.Cheatgrass has increased fire fre- dustbowl,has substantially lower allocation to roots compared with quency by a factor of more than ten in the >40 million hectares native prairie grasses.Soil under A.cristatum has lower levels of (1ha=10'm2)thatit now dominates2. available nitrogen and-25%less total carbon than native prairie soil, Species-induced changes in microclimate can be just as impor- so the introduction of this species resulted in an equivalent reduction tant as the direct impacts of environmental change.For example,in of 480 x 102g carbon stored in soils7.Soil invertebrates,such as late-successional boreal forests,where soil temperatures have a earthworms and termites,also alter turnover of organic matter and strong influenceon nutrient supplyand productivity,the presence of nutrient supply,thereby influencing the species composition of the moss,which reduces heat flux into the soil,contributes to the stability aboveground flora and fauna's of permafrost(frozen soils)and the characteristically low rates of Species can also influence disturbance regime.For example, nutrient cycling".As fire frequency increases in response to high-lat- several species of nutritious but flammable grasses were introduced itude warming,moss biomass declines,permafrost becomes less sta- to the Hawaiian Islands to support cattle grazing.Some of these ble,the nutrient supply increases,and the species composition of grasses spread into protected woodlands,where they caused a 300- forests is altered.Plant traits can also influence climate at larger fold increase in the extent of fire.Most ofthe woody plants,including scales.Simulations with general circulation models indicate that some endangered species,are eliminated by fire,whereas grasses widespread replacement of deep-rooted tropical trees by shallow- Figure 3 Scenarios of change in species diversity in selected biomes by the year 2100.The values are the projected change in diversity for each □Other ☐Land use biome relative to the biome with greatest projected diversity change. Exotic Climate Biomes are:tropical forests (T),grasslands (G),Mediterranean (M), desert (D).north temperate forests (N).boreal forests (B)and arctic(A). Projected change in species diversity is calculated assuming three altemnative scenarios of interactions among the causes of diversity change.Scenario 1 assumes no interaction among causes of diversity change,so that the total change in diversity is the sum of the changes caused by each driver of diversity change.Scenario 2 assumes that only the factor with the greatest impact on diversity influences diversity change.Scenario 3 assumes that factors causing change in biodiversity interact multiplicatively to determine diversity change.For scenarios 1 and 2,we show the relative importance of the major causes of projected change in diversity.These causes are climatic change,change in land 0.4 use,introduction of exotic species,and changes in atmospheric Co. and/or nitrogen deposition (labelled 'other).The graph shows that all biomes are projected to experience substantial change in species diversity by 2100,that the most important causes of diversity change differ among biomes,and that the patterns of diversity change depend on assumptions about the nature of interactions among the causes of diversity change.Projected biodiversity change is most similar among TGM D N B G M D N B biomes if causes of diversity change do not interact(scenario 1)and Scenario 1 Scenario 2 Scenario 3 differ most srongy among biomes thecauses of biodiversity change interact multiplicatively (scenario 3). 236 2000 Macmillan Magazines Ltd NATURE VOL 40511 MAY 2000 www.nature.comaccessed by vegetation, enhanced productivity, and increased surface litter and salts. This inhibited the regeneration of many native species, leading to a general reduction in biodiversity16. The perennial tussock grass, Agropyron cristatum, which was widely introduced to the northern Great Plains of North America after the 1930s ‘dustbowl’, has substantially lower allocation to roots compared with native prairie grasses. Soil under A. cristatum has lower levels of available nitrogen and ~25% less total carbon than native prairie soil, so the introduction of this species resulted in an equivalent reduction of 480 2 1012 g carbon stored in soils17. Soil invertebrates, such as earthworms and termites, also alter turnover of organic matter and nutrient supply, thereby influencing the species composition of the aboveground flora and fauna18. Species can also influence disturbance regime. For example, several species of nutritious but flammable grasses were introduced to the Hawaiian Islands to support cattle grazing. Some of these grasses spread into protected woodlands, where they caused a 300- fold increase in the extent of fire. Most of the woody plants, including some endangered species, are eliminated by fire, whereas grasses rebound quickly19. Similar increases in the ecological role of fire resulting from grass invasions have been widely observed in the Americas, Australia and elsewhere in Oceania. The invasion of cheatgrass (Bromus tectorum) into western North America is one of the most extensive of these invasions. Cheatgrass has increased fire frequency by a factor of more than ten in the >40 million hectares (1 ha = 104 m2 ) that it now dominates20. Species-induced changes in microclimate can be just as important as the direct impacts of environmental change. For example, in late-successional boreal forests, where soil temperatures have a strong influence on nutrient supply and productivity, the presence of moss, which reduces heat flux into the soil, contributes to the stability of permafrost (frozen soils) and the characteristically low rates of nutrient cycling21. As fire frequency increases in response to high-latitude warming, moss biomass declines, permafrost becomes less stable, the nutrient supply increases, and the species composition of forests is altered. Plant traits can also influence climate at larger scales. Simulations with general circulation models indicate that widespread replacement of deep-rooted tropical trees by shallowinsight review articles 236 NATURE | VOL 405 | 11 MAY 2000 | www.nature.com Figure 3 Scenarios of change in species diversity in selected biomes by the year 2100. The values are the projected change in diversity for each biome relative to the biome with greatest projected diversity change6 . Biomes are: tropical forests (T), grasslands (G), Mediterranean (M), desert (D), north temperate forests (N), boreal forests (B) and arctic (A). Projected change in species diversity is calculated assuming three alternative scenarios of interactions among the causes of diversity change. Scenario 1 assumes no interaction among causes of diversity change, so that the total change in diversity is the sum of the changes caused by each driver of diversity change. Scenario 2 assumes that only the factor with the greatest impact on diversity influences diversity change. Scenario 3 assumes that factors causing change in biodiversity interact multiplicatively to determine diversity change. For scenarios 1 and 2, we show the relative importance of the major causes of projected change in diversity. These causes are climatic change, change in land use, introduction of exotic species, and changes in atmospheric CO2 and/or nitrogen deposition (labelled ‘other’). The graph shows that all biomes are projected to experience substantial change in species diversity by 2100, that the most important causes of diversity change differ among biomes, and that the patterns of diversity change depend on assumptions about the nature of interactions among the causes of diversity change. Projected biodiversity change is most similar among biomes if causes of diversity change do not interact (scenario 1) and differ most strongly among biomes if the causes of biodiversity change interact multiplicatively (scenario 3). 1 0.8 0.6 0.4 0.2 0 TGMDNBA TGMDNBA TGMDNBA Relative diversity change (proportion of maximum) Scenario 1 Scenario 2 Scenario 3 Other Exotic Land use Climate There has been substantial debate over both the form of the relationship between species richness and ecosystem processes and the mechanisms underlying these relationships85. Theoretically, rates of ecosystem processes might increase linearly with species richness if all species contribute substantially and in unique ways to a given process — that is, have complementary niches. This relationship is likely to saturate as niche overlap, or ‘redundancy’, increases at higher levels of diversity86. Several experiments indicate such an asymptotic relationship of ecosystem process rates with species richness. An asymptotic relationship between richness and process rates could, however, arise from a ‘sampling effect’ of increased probability of including a species with strong ecosystem effects, as species richness increases13. The sampling effect has at least two interpretations. It might be an important biological property of communities that influences process rates in natural ecosystems13, or it might be an artefact of species-richness experiments in which species are randomly assigned to treatments, rather than following community assembly rules that might occur in nature87. Finally, ecosystem process rates may show no simple correlation with species richness. However, the lack of a simple statistical relationship between species richness and an ecosystem process may mask important functional relationships. This could occur, for example, if process rates depend strongly on the traits of certain species or if species interactions determine the species traits that are expressed (the ‘idiosyncratic hypothesis’)7 . This mechanistic debate is important scientifically for understanding the functioning of ecosystems and effective management of their biotic resources. Regardless of the outcome of the debate, conserving biodiversity is essential because we rarely know a priori which species are critical to current functioning or provide resilience and resistance to environmental changes. Box 1 Species richness and ecosystem functioning © 2000 Macmillan Magazines Ltd