正在加载图片...
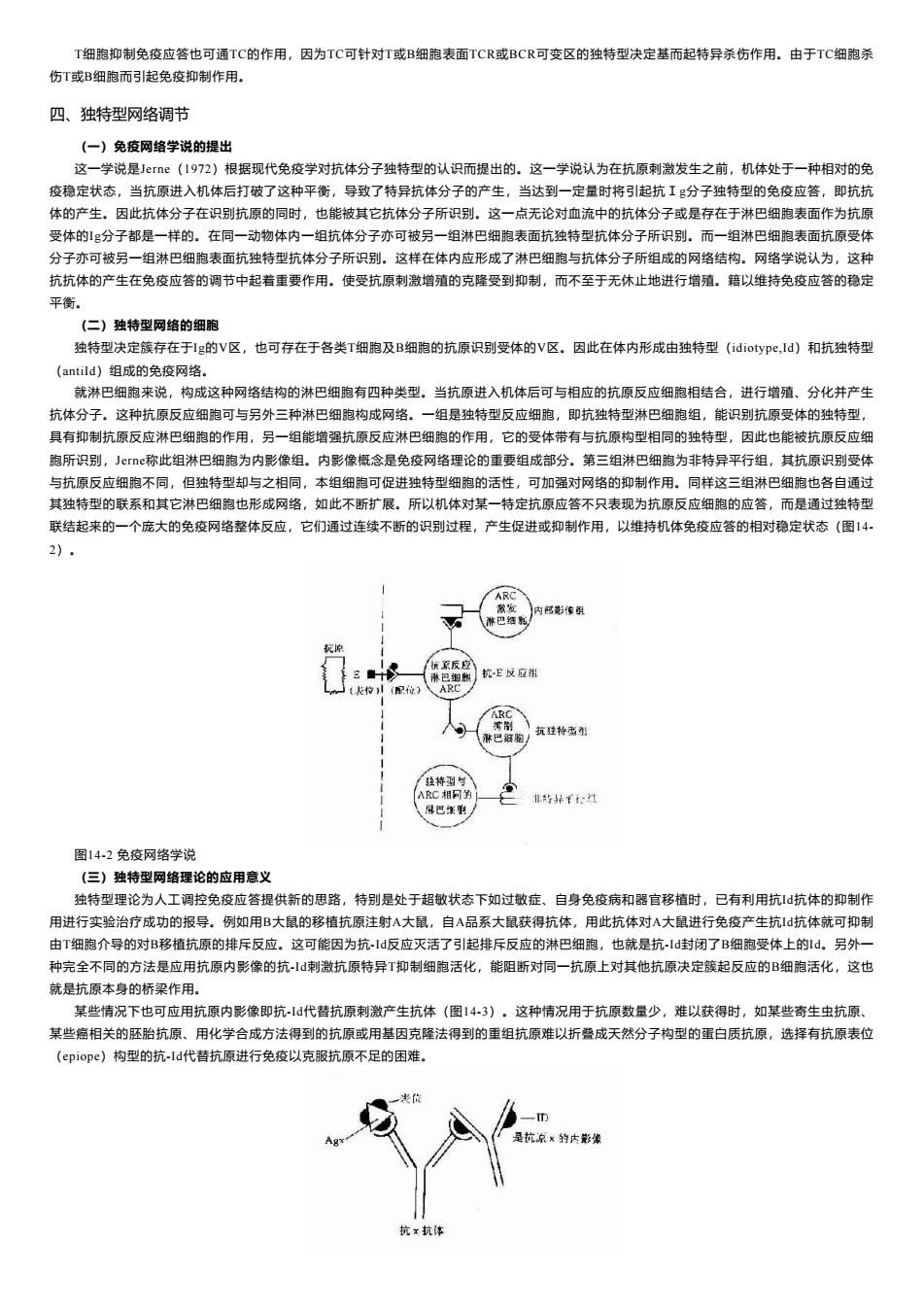
T细胞抑制免疫应答也可通TC的作用。因为TC可针对T或B细孢表面TCR或BCR可变区的独特型决定基而起特异杀伤作用。由于TC细胞杀 伤T或B细胞而引起免疫抑制作用, 四、独特型网络调节 (一)免废网络学说的提出 这一学说是ee(1972)根晷现代免疫学对抗体分子独特型的认识而提出的。这一学说认为在抗原刺微发生之前,机体处于一种相对的免 疫稳定状态,当抗原进入机体后打破了这种平衡,导致了特异抗体分子的产生,当达到一定量时将引起抗Ig分子独特型的免疫应答,即抗抗 体的产生。因此抗体分子在识到抗原的同时,也能被其它抗体分子所识别。这一点无论对血流中的抗体分子或是存在于淋巴细跑表面作为抗原 受体的1g分子都是一样的。在同一动物体内一组抗体分子亦可被另一组淋巴细胞表面抗独特型抗体分子所识别。而一组淋巴细胞表面抗原受体 分子亦可被另一组淋巴细胞表面抗独特型抗体分子所识别。这样在体内应形成了淋巴细胞与抗体分子所组成的网络结构。网络学说认为,这种 抗抗体的产生在免液应答的调节中起着重要作用,使受抗原刺激增殖的克降受到和制,而不至于无休止地进行增殖。籍以维持免疫应答的稳定 平衡。 (二)独特型网格的细胞 独特型决定簇存在于刊g的V区,也可存在于各类T细胞及B细跑的抗原识别受体的V区。因此在体内形成由独特型(dioype,ld)和抗独特型 antild)组成的免疮网 就淋巴细泡来说,构成这种网络结物的淋巴细胞有四种类型。当抗原进入机体后可与相应的抗原反应细胞相结合,进行增殖、分化并产生 抗体分子。这种抗原反应细胞可与另外三种淋巴细构成风络。一组是独特型反应细胞。即抗独特型淋巴细胞组,能识别抗原受体的独特型 有抑制抗原反应淋巴细胞的作用, 组能强抗原反应巴细胞的作用它的受体带有与抗原构型相同的独特型因此也能被抗原反应组 所识别,ee称此组淋巴细胞为内影像组。内影像概念是免疫网络理论的重要组成部分。第三组 细胞为非导平行组其抗原识到受女 与抗反应细的 组细可促讲独结 细电的应答,而是通过独特无 联结起来的一个底大的免疫网络整体反应,它们通过连续不断的识别过程,产生促进或抑制作用,以维持机体免疫应答的相对稳定状态(图1 2 人x 品 。非5好仁江 图14.2免疫网络学说 (三)独特型网络理论的应用意义 独特型理论为人工调控免疫应答提供新的思路,特别是处于超敏状态下如过敏在,自身免疫病和器官移植时,已有利用抗1抗体的抑制作 用进行实验治疗成功的报导,例如用B大鼠的移植抗原注射八大鼠,自A品系大鼠获得抗体,用此抗体对A大鼠进行免疫产生抗1抗体就可抑制 由T细胞介导的对B移植抗原的排斥反应。这可能因为抗ld反应灭活了引起排拆反应的淋巴细孢,也就是抗,1封闭了B细胞受体上的d,另外 种完全不同的方法是应用抗原内影像的抗I刺激抗原特异T抑制细胞活化,能阻断对同一抗原上对其他抗原决定簇起反应的B细胞活化,这也 就是抗原本身的桥梁作用 某些情况下也可应用抗原内影像即抗-d代善抗原刺激产生抗体(图143),这种情况用于抗原数量少,难以获得时,如某些奇生虫抗原, 某些癌相关的胚胎抗原。用化学合成方法得到的抗原或用基因克降法得到的重组抗原难以折叠成天然分子构型的蛋白质抗原,选择有抗原表位 (epiope)构型的抗-ld代替抗原进行免疫以克服抗原不足的因难.T细胞抑制免疫应答也可通TC的作用,因为TC可针对T或B细胞表面TCR或BCR可变区的独特型决定基而起特异杀伤作用。由于TC细胞杀 伤T或B细胞而引起免疫抑制作用。 四、独特型网络调节 (一)免疫网络学说的提出 这一学说是Jerne(1972)根据现代免疫学对抗体分子独特型的认识而提出的。这一学说认为在抗原刺激发生之前,机体处于一种相对的免 疫稳定状态,当抗原进入机体后打破了这种平衡,导致了特异抗体分子的产生,当达到一定量时将引起抗Ig分子独特型的免疫应答,即抗抗 体的产生。因此抗体分子在识别抗原的同时,也能被其它抗体分子所识别。这一点无论对血流中的抗体分子或是存在于淋巴细胞表面作为抗原 受体的Ig分子都是一样的。在同一动物体内一组抗体分子亦可被另一组淋巴细胞表面抗独特型抗体分子所识别。而一组淋巴细胞表面抗原受体 分子亦可被另一组淋巴细胞表面抗独特型抗体分子所识别。这样在体内应形成了淋巴细胞与抗体分子所组成的网络结构。网络学说认为,这种 抗抗体的产生在免疫应答的调节中起着重要作用。使受抗原刺激增殖的克隆受到抑制,而不至于无休止地进行增殖。籍以维持免疫应答的稳定 平衡。 (二)独特型网络的细胞 独特型决定簇存在于Ig的V区,也可存在于各类T细胞及B细胞的抗原识别受体的V区。因此在体内形成由独特型(idiotype,Id)和抗独特型 (antiId)组成的免疫网络。 就淋巴细胞来说,构成这种网络结构的淋巴细胞有四种类型。当抗原进入机体后可与相应的抗原反应细胞相结合,进行增殖、分化并产生 抗体分子。这种抗原反应细胞可与另外三种淋巴细胞构成网络。一组是独特型反应细胞,即抗独特型淋巴细胞组,能识别抗原受体的独特型, 具有抑制抗原反应淋巴细胞的作用,另一组能增强抗原反应淋巴细胞的作用,它的受体带有与抗原构型相同的独特型,因此也能被抗原反应细 胞所识别,Jerne称此组淋巴细胞为内影像组。内影像概念是免疫网络理论的重要组成部分。第三组淋巴细胞为非特异平行组,其抗原识别受体 与抗原反应细胞不同,但独特型却与之相同,本组细胞可促进独特型细胞的活性,可加强对网络的抑制作用。同样这三组淋巴细胞也各自通过 其独特型的联系和其它淋巴细胞也形成网络,如此不断扩展。所以机体对某一特定抗原应答不只表现为抗原反应细胞的应答,而是通过独特型 联结起来的一个庞大的免疫网络整体反应,它们通过连续不断的识别过程,产生促进或抑制作用,以维持机体免疫应答的相对稳定状态(图14- 2)。 图14-2 免疫网络学说 (三)独特型网络理论的应用意义 独特型理论为人工调控免疫应答提供新的思路,特别是处于超敏状态下如过敏症、自身免疫病和器官移植时,已有利用抗Id抗体的抑制作 用进行实验治疗成功的报导。例如用B大鼠的移植抗原注射A大鼠,自A品系大鼠获得抗体,用此抗体对A大鼠进行免疫产生抗Id抗体就可抑制 由T细胞介导的对B移植抗原的排斥反应。这可能因为抗-Id反应灭活了引起排斥反应的淋巴细胞,也就是抗-Id封闭了B细胞受体上的Id。另外一 种完全不同的方法是应用抗原内影像的抗-Id刺激抗原特异T抑制细胞活化,能阻断对同一抗原上对其他抗原决定簇起反应的B细胞活化,这也 就是抗原本身的桥梁作用。 某些情况下也可应用抗原内影像即抗-Id代替抗原刺激产生抗体(图14-3)。这种情况用于抗原数量少,难以获得时,如某些寄生虫抗原、 某些癌相关的胚胎抗原、用化学合成方法得到的抗原或用基因克隆法得到的重组抗原难以折叠成天然分子构型的蛋白质抗原,选择有抗原表位 (epiope)构型的抗-Id代替抗原进行免疫以克服抗原不足的困难