正在加载图片...
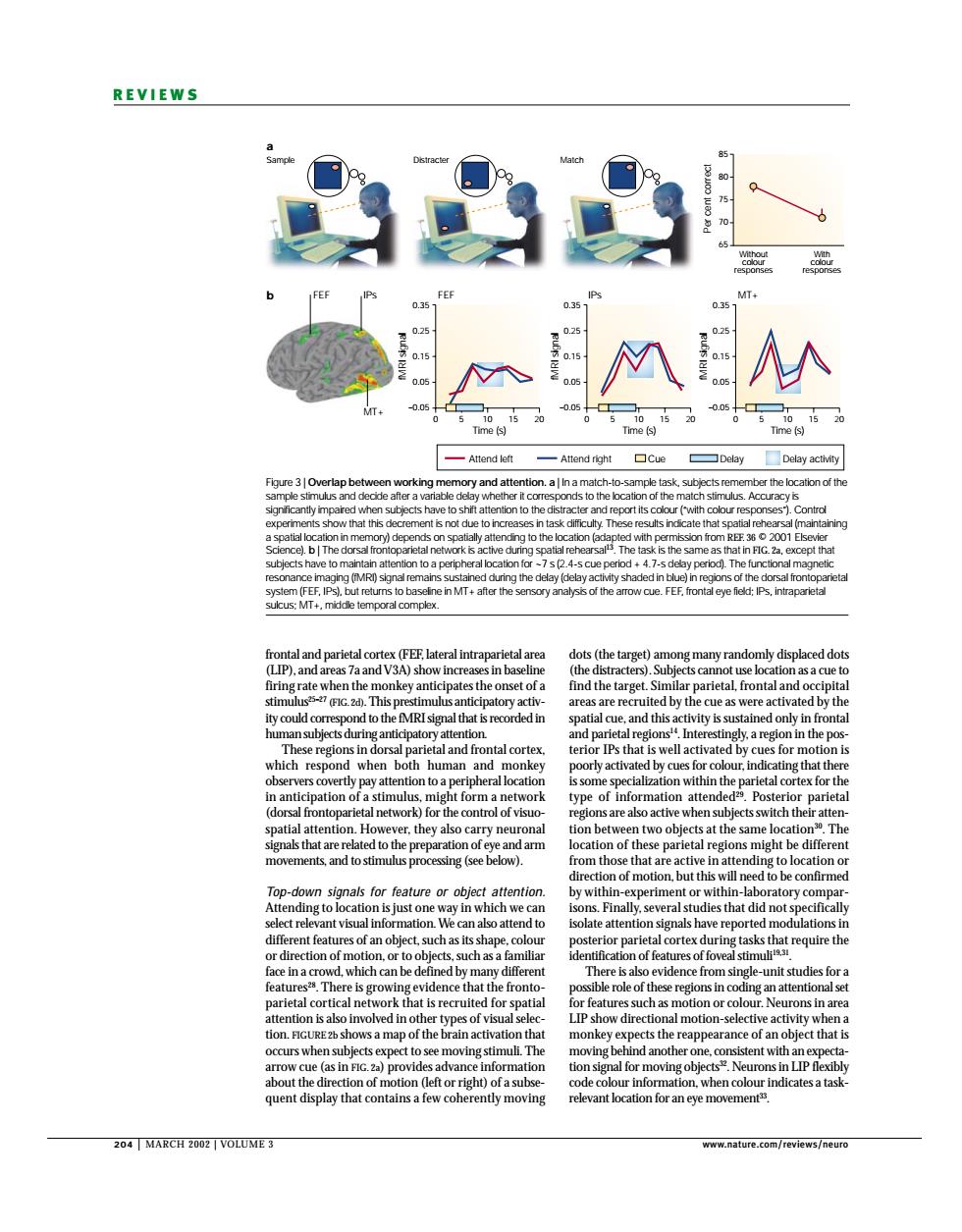
REVIEWS M Time (s) me is Attend loft Attend right the location of the V3 in b his p which res human nantiei pation o a sti patial attention r the biects at th Th down signals for feature or obied attentio n-lab nt visu 01 or pari face in The that the the reappe f an object that quent display that contains a few cohe ently moving relevant location for an eye movement 204 MARCH2002 VOLUM正3 204 | MARCH 2002 | VOLUME 3 www.nature.com/reviews/neuro REVIEWS dots (the target) among many randomly displaced dots (the distracters). Subjects cannot use location as a cue to find the target. Similar parietal, frontal and occipital areas are recruited by the cue as were activated by the spatial cue, and this activity is sustained only in frontal and parietal regions14. Interestingly, a region in the posterior IPs that is well activated by cues for motion is poorly activated by cues for colour, indicating that there is some specialization within the parietal cortex for the type of information attended29. Posterior parietal regions are also active when subjects switch their attention between two objects at the same location30. The location of these parietal regions might be different from those that are active in attending to location or direction of motion, but this will need to be confirmed by within-experiment or within-laboratory comparisons. Finally, several studies that did not specifically isolate attention signals have reported modulations in posterior parietal cortex during tasks that require the identification of features of foveal stimuli19,31. There is also evidence from single-unit studies for a possible role of these regions in coding an attentional set for features such as motion or colour. Neurons in area LIP show directional motion-selective activity when a monkey expects the reappearance of an object that is moving behind another one, consistent with an expectation signal for moving objects32. Neurons in LIP flexibly code colour information, when colour indicates a taskrelevant location for an eye movement33. frontal and parietal cortex (FEF, lateral intraparietal area (LIP), and areas 7a and V3A) show increases in baseline firing rate when the monkey anticipates the onset of a stimulus25–27 (FIG. 2d). This prestimulus anticipatory activity could correspond to the fMRI signal that is recorded in human subjects during anticipatory attention. These regions in dorsal parietal and frontal cortex, which respond when both human and monkey observers covertly pay attention to a peripheral location in anticipation of a stimulus, might form a network (dorsal frontoparietal network) for the control of visuospatial attention. However, they also carry neuronal signals that are related to the preparation of eye and arm movements, and to stimulus processing (see below). Top-down signals for feature or object attention. Attending to location is just one way in which we can select relevant visual information. We can also attend to different features of an object, such as its shape, colour or direction of motion, or to objects, such as a familiar face in a crowd, which can be defined by many different features28. There is growing evidence that the frontoparietal cortical network that is recruited for spatial attention is also involved in other types of visual selection. FIGURE 2b shows a map of the brain activation that occurs when subjects expect to see moving stimuli. The arrow cue (as in FIG. 2a) provides advance information about the direction of motion (left or right) of a subsequent display that contains a few coherently moving Sample a b Distracter Match 85 80 75 70 65 Per cent correct Without colour responses With colour responses 0.35 0.25 0.15 0.05 –0.05 0 5 10 FEF IPs IPs FEF 15 20 0.35 0.25 0.15 0.05 –0.05 0 5 10 MT+ Time (s) Time (s) Time (s) fMRI signal fMRI signal fMRI signal 15 20 0.35 0.25 0.15 0.05 –0.05 0 5 10 15 20 Attend left Delay activity Attend right Cue Delay MT+ Figure 3 | Overlap between working memory and attention. a | In a match-to-sample task, subjects remember the location of the sample stimulus and decide after a variable delay whether it corresponds to the location of the match stimulus. Accuracy is significantly impaired when subjects have to shift attention to the distracter and report its colour (“with colour responses”). Control experiments show that this decrement is not due to increases in task difficulty. These results indicate that spatial rehearsal (maintaining a spatial location in memory) depends on spatially attending to the location (adapted with permission from REF. 36 © 2001 Elsevier Science). b | The dorsal frontoparietal network is active during spatial rehearsal13. The task is the same as that in FIG. 2a, except that subjects have to maintain attention to a peripheral location for ~7 s (2.4-s cue period + 4.7-s delay period). The functional magnetic resonance imaging (fMRI) signal remains sustained during the delay (delay activity shaded in blue) in regions of the dorsal frontoparietal system (FEF, IPs), but returns to baseline in MT+ after the sensory analysis of the arrow cue. FEF, frontal eye field; IPs, intraparietal sulcus; MT+, middle temporal complex