正在加载图片...
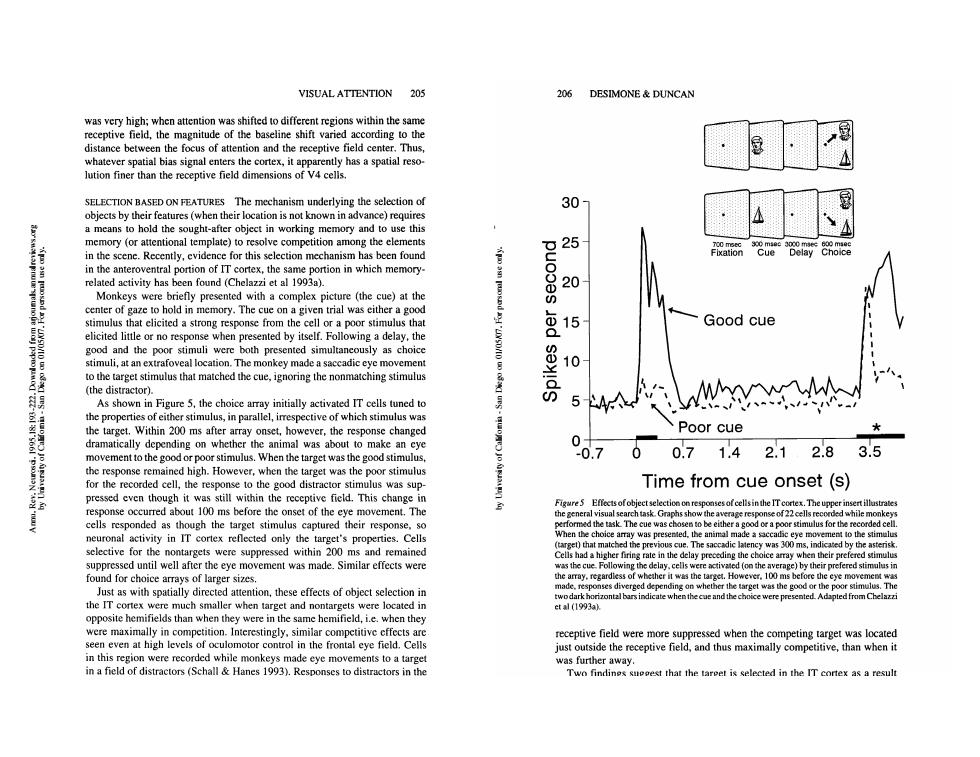
VISUALATTENTION 205 206 DESIMONE DUNCAN receptive field,the magnitude of the e shift varied ccording to the whatever spatial bias signal enters the cortex,it apparently has a spatial reso- lution finer than the receptive field dimensions of v4 celis. SELECTION BASED ON FEATURES The mechanism underlying the selection of 30 objects by their features(when their location is not known in advance)requires a means to hold the sought-after object in working memory and to use this memory (or attentional template)to resolve competition among the elements in the scene.Recently,evidence for this selection mechanism has been found 25 in the anteroventral portion of IT cortex,the same portion in which memory related activity has been found (Chelazzi et al 1993a) briefly pre rotgactoho esented with a co mp ex picture (the cue)at the cent ory.The cue on a goo at elicited a strong espons Good cue icited or no resp onse elf.Fol ⑧15 ga delay,the and the po oth presented nuli,at an extrafoveal lo tion.The monkey made a sacc to the target stimulus that matched the cue,ignoring the nonmatching stimulus (the distractor). As shown in Figure 5.the choice array initially activated IT cells tuned to 小 the properties of either stimulus,in parallel,irrespective of which stimulus was the target.Within 200 ms after array onset,however,the response changed Poor cue dramatically depending on whether the animal was about to make an cye 0 movement to the good or poor stimulus.When the target was the good stimulus .7 00.71.42.12.83.5 the response remained high.However.when the tareet was the poor stimulus for the recorded cell,the resp onse to the good dist actor stimulus was sup Time from cue onset (s) pressed even tho gh ithin ms be ve field.This change Figure 5 Effects ofobject selection onr 100 ocellsinthe ITcore.The upp movemen he general aphs sho erage r the ta ity in IT cortex reflected only th target's prop Whenthe choice animal made a sacc m to the stin and remained ecedighcchaieea suppressed until well after the eye movement was made.Similar effects were the cu ving the delay.celis found for choice arrays of larger sizes. n whe Just as with spatially directed attention,these effects of object selection in s diverged pending o get was the good or th the IT cortex were much smaller when target and nontargets were located in et al (1993ak opposite hemifields than when they were in the same hemifield,i.e.when they were maximally in competition.Interestingly,similar competitive effects are receptive field were more suppressed when the competing target was located seen even at high levels of oculomotor control in the frontal eye field.Cells just ouside the receptive field,and thus maximally competitive,than when it in this region were recorded while monkeys made eye movements to a target was further in a field of distractors(Schall Hanes 1993).Responses to distractors in the Two find sueeest that the taroet is selected in the IT cortex as a result VISUAL ATTENTION 205 was very high; when attention was shifted to different regions within the same receptive field, the magnitude of the baseline shift varied according to the distance between the focus of attention and the receptive field center. Thus, whatever spatial bias signal enters the cortex, it apparently has a spatial resolution finer than the receptive field dimensions of V4 cells. SELECTION BASED ON FEATURES The mechanism underlying the selection of objects by their features (when their location is not known in advance) requires a means to hold the sought-after object in working memory and to use this memory (or attentional template) to resolve competition among the elements in the scene. Recently, evidence for this selection mechanism has been found in the anteroventral portion of IT cortex, the same portion in which memoryrelated activity has been found (Chelazzi et al 1993a). Monkeys were briefly presented with a complex picture (the cue) at the center of gaze to hold in memory. The cue on a given trial was either a good stimulus that elicited a strong response from the cell or a poor stimulus that elicited little or no response when presented by itself. Following a delay, the good and the poor stimuli were both presented simultaneously as choice stimuli, at an extrafoveal location. The monkey made a saccadic eye movement to the target stimulus that matched the cue, ignoring the nonmatching stimulus (the distractor). As shown in Figure 5, the choice array initially activated IT cells tuned to the properties of either stimulus, in parallel, irrespective of which stimulus was the target. Within 200 ms after array onset, however, the response changed dramatically depending on whether the animal was about to make an eye movement to the good or poor stimulus. When the target was the good stimulus, the response remained high. However, when the target was the poor stimulus for the recorded cell, the response to the good distractor stimulus was suppressed even though it was still within the receptive field. This change in response occurred about 100 ms before the onset of the eye movement. The cells responded as though the target stimulus captured their response, so neuronal activity in IT cortex reflected only the target’s properties. Cells selective for the nontargets were suppressed within 200 ms and remained suppressed until well after the eye movement was made. Similar effects were found for choice arrays of larger sizes. Just as with spatially directed attention, these effects of object selection in the IT cortex were much smaller when target and nontargets were located in opposite hemifields than when they were in the same hemifield, i.e. when they were maximally in competition. Interestingly, similar competitive effects are seen even at high levels of oculomotor control in the frontal eye field. Cells in this region were recorded while monkeys made eye movements to a target in a field of distractors (Schall & Hanes 1993). Responses to distractors in the www.annualreviews.org/aronline Annual Reviews Annu. Rev. Neurosci. 1995.18:193-222. Downloaded from arjournals.annualreviews.org by University of California - San Diego on 01/05/07. For personal use only. 206 DESIMONE & DUNCAN 30- 25- 20- ~15- ,., ~ -- Poor cue ,,-~, -0.7 0 0.7 1.4 2.1 2.8 3.5 Time from cue onset (s) Figure5 Effects ofobject selection on responses of cells in the IT cortex. The upper insert illustrates the general visual search task. Graphshow the average response of 22 cells recorded while monkeys performed the task. The cue was chosen to be either a good or a poor stimulus for the recorded cell. When the choice array was presented, the animal made a saccadic eye movement to the stimulus (target) that matched the previous cue. The saccadic latency was 300 ms, indicated by the asterisk. Cells had a higher firing rate in the delay preceding the choice array when their prefered stimulus was the cue. Following the delay, cells were activated (on the average) by their prefered stimulus the array, regardless of whether it was the target. However, 100 ms before the eye movement was made, responses diverged depending on whether the target was the good or the poor stimulus. The two dark horizontal bars indicate when the cue and the choice were presented. Adapted from Chelazzi et al (1993a). receptive field were more suppressed when the competing target was located just outside the receptive field, and thus maximally competitive, than when it was further away. Two findings suggest that the target is selected in the IT cortex as a result www.annualreviews.org/aronline Annual Reviews Annu. Rev. Neurosci. 1995.18:193-222. Downloaded from arjournals.annualreviews.org by University of California - San Diego on 01/05/07. For personal use only