正在加载图片...
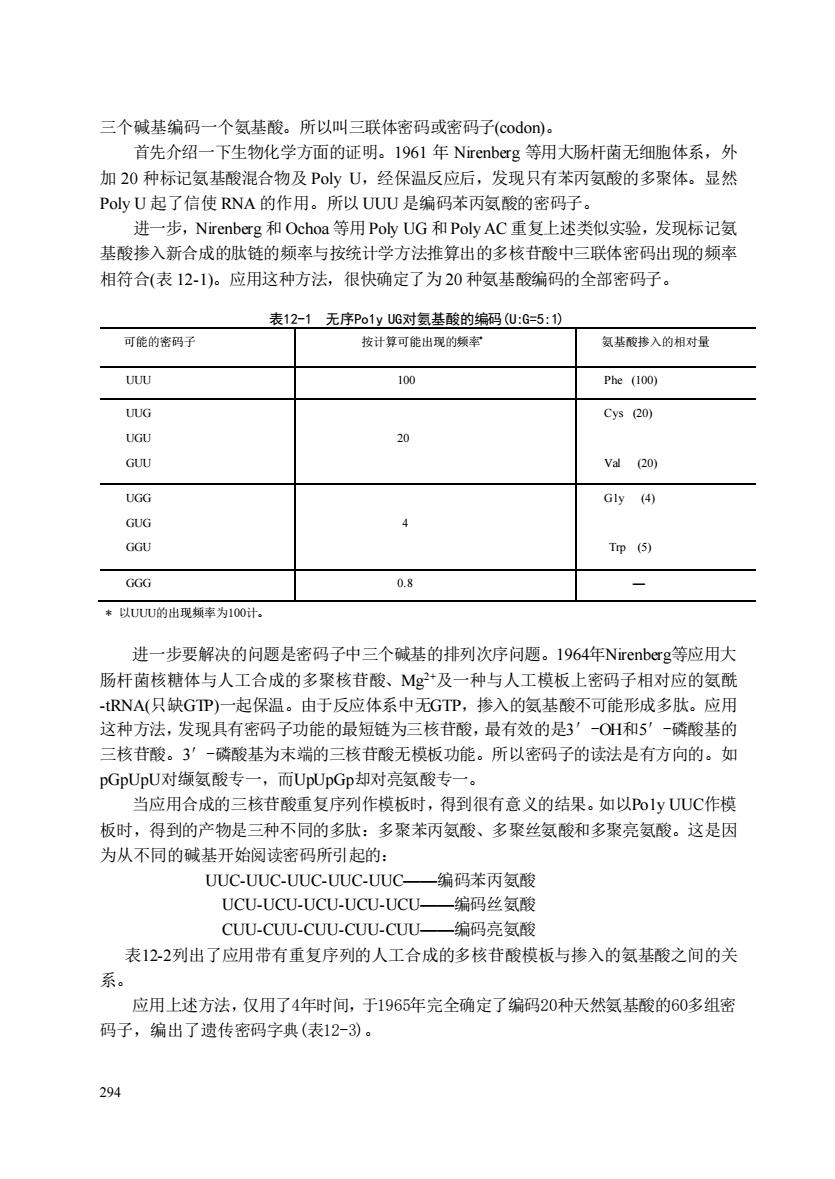
三个碱基编码一个氨基酸。所以叫三联体密码或密码子(codon))。 首先介绍一下生物化学方面的证明。1961年Nirenberg等用大肠杆菌无细胞体系,外 加20种标记氨基酸混合物及PyU,经保温反应后,发现只有苯丙氨酸的多聚体。显然 PoyU起了信使RNA的作用。所以UUU是编码苯丙氨酸的密码子。 进一步,Nirenberg和Ochoa等用PolyUG和Poly AC重复上述类似实验,发现标记氨 基酸掺入新合成的肤链的频率与按统计学方法推算出的多核苷酸中三联体密码出现的颜率 相符合(表12-1)。应用这种方法,很快确定了为20种氨基酸编码的全部密码子。 表12-1无序Po1yUG对氨基酸的编码U:G=5:1) 可能的密码子 按计算可能出现的频率 氨基酸摻入的相对量 UUU Phe (100) UUG Cys (20) 20 GUU va(20) UGG Gy④ GUG GGU Trp (5) GGG 08 *以UUU的出现频率为100计. 进一步要解决的问题是密码子中三个碱基的排列次序问题。1964年Nirenberg等应用大 肠杆菌核糖体与人工合成的多聚核苷酸、Mg2+及一种与人工模板上密码子相对应的氨酰 -RNA(只缺GP)一起保温。由于反应体系中无GTP,掺入的奥基酸不可能形成多肽。应用 这种方法,发现具有密码子功能的最短链为三核苷酸,最有效的是3'0H和5'-磷酸基的 三核苷酸。3'一磷酸基为末端的三核苷酸无模板功能。所以密码子的读法是有方向的。如 pGpUpU对缬氨酸专一,而UpUpGp却对亮氨酸专一。 当应用合成的三核苷酸重复序列作模板时,得到很有意义的结果。如以Po1yUUC作模 板时,得到的产物是三种不同的多肽:多聚苯丙氨酸、多聚丝氨酸和多聚亮氨酸。这是因 为从不同的碱基开始阅读密码所引起的: UUC-UUC-UUC-UUC-UUC- 编码苯丙氨酸 UCU-UCU-UCU-UCU-UCU- -编码丝氨酸 CUU-CUU-CUU-CUU-CUU一编码亮氨酸 表122列出了应用带有重复序列的人工合成的多核苷酸模板与掺入的氨基酸之间的关 系。 应用上述方法,仅用了4年时间,于1965年完全确定了编码20种天然氨基酸的60多组密 码子,编出了遗传密码字典(表12-)。 294 294 三个碱基编码一个氨基酸。所以叫三联体密码或密码子(codon)。 首先介绍一下生物化学方面的证明。1961 年 Nirenberg 等用大肠杆菌无细胞体系,外 加 20 种标记氨基酸混合物及 Poly U,经保温反应后,发现只有苯丙氨酸的多聚体。显然 Poly U 起了信使 RNA 的作用。所以 UUU 是编码苯丙氨酸的密码子。 进一步,Nirenberg 和 Ochoa 等用Poly UG 和Poly AC 重复上述类似实验,发现标记氨 基酸掺入新合成的肽链的频率与按统计学方法推算出的多核苷酸中三联体密码出现的频率 相符合(表 12-1)。应用这种方法,很快确定了为 20 种氨基酸编码的全部密码子。 表12-1 无序Po1y UG对氨基酸的编码(U:G=5:1) 可能的密码子 按计算可能出现的频率* 氨基酸掺入的相对量 UUU 100 Phe (100) UUG UGU GUU 20 Cys (20) Val (20) UGG GUG GGU 4 G1y (4) Trp (5) GGG 0.8 — * 以UUU的出现频率为100计。 进一步要解决的问题是密码子中三个碱基的排列次序问题。1964年Nirenberg等应用大 肠杆菌核糖体与人工合成的多聚核苷酸、Mg2+及一种与人工模板上密码子相对应的氨酰 -tRNA(只缺GTP)一起保温。由于反应体系中无GTP,掺入的氨基酸不可能形成多肽。应用 这种方法,发现具有密码子功能的最短链为三核苷酸,最有效的是3′-OH和5′-磷酸基的 三核苷酸。3′-磷酸基为末端的三核苷酸无模板功能。所以密码子的读法是有方向的。如 pGpUpU对缬氨酸专一,而UpUpGp却对亮氨酸专一。 当应用合成的三核苷酸重复序列作模板时,得到很有意义的结果。如以Po1y UUC作模 板时,得到的产物是三种不同的多肽:多聚苯丙氨酸、多聚丝氨酸和多聚亮氨酸。这是因 为从不同的碱基开始阅读密码所引起的: UUC-UUC-UUC-UUC-UUC——编码苯丙氨酸 UCU-UCU-UCU-UCU-UCU——编码丝氨酸 CUU-CUU-CUU-CUU-CUU——编码亮氨酸 表12-2列出了应用带有重复序列的人工合成的多核苷酸模板与掺入的氨基酸之间的关 系。 应用上述方法,仅用了4年时间,于1965年完全确定了编码20种天然氨基酸的60多组密 码子,编出了遗传密码字典(表12-3)