正在加载图片...
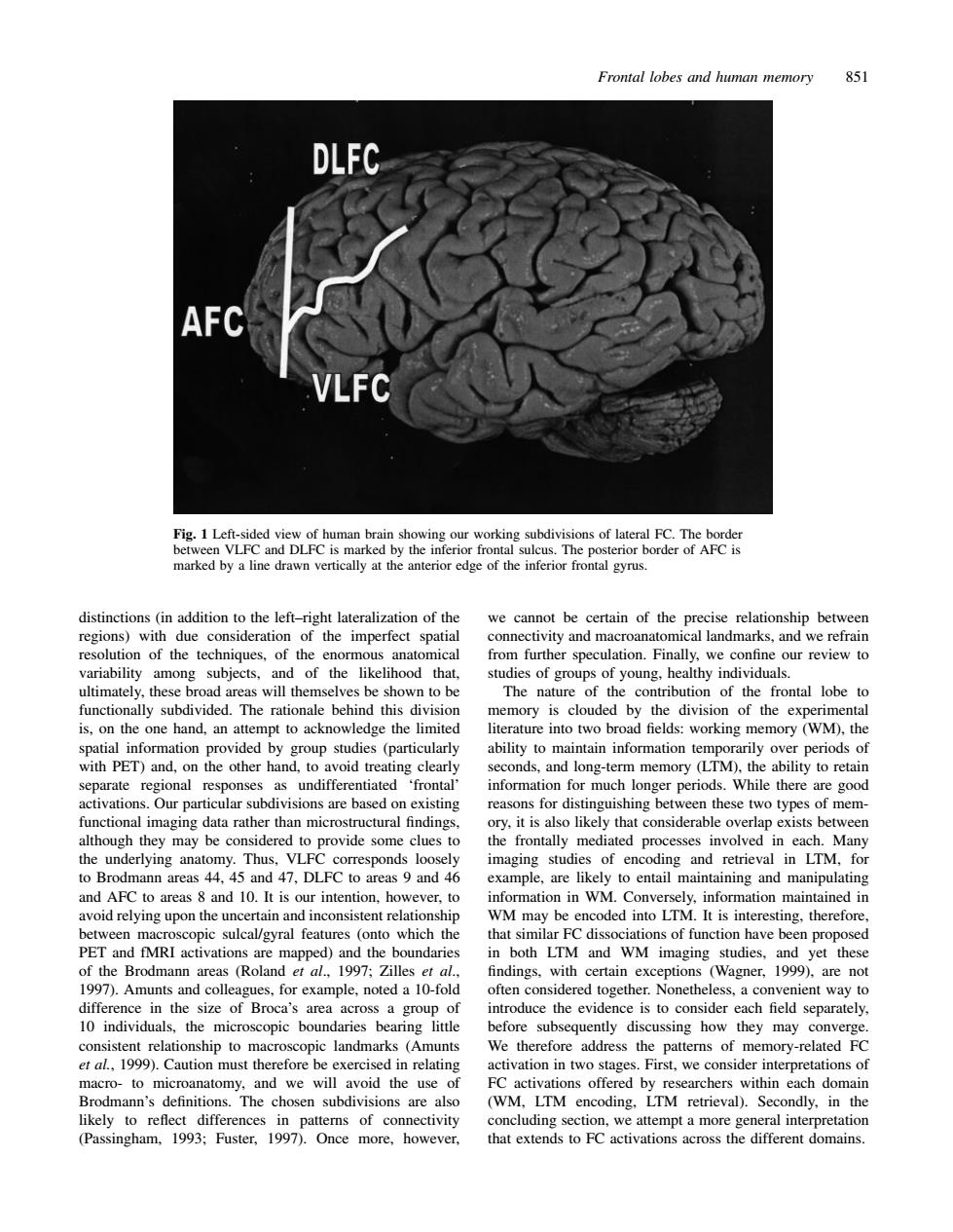
Frontal lobes and human memory 851 DLFC AFC Fig.1 Le ving our working subdivi marked by a ine drawn vertically at the anterior edge of the inferior frontal gyrus. distinctions (in addition to the left-right lateralization of the we cannot be certain of the precise relationship between al landma am subiects and of the likelihood that ultimately,these broad areas will themselves be shown to be The nature of the contribution of the frontal lobe to ally subdivided heaona behind thi ory is cloude d by the divis of the exp inf to ack ntain info nds,and long-term memory (LTM),the ability to retain separate regional 'frontal information for much longer periods.While there are good Our d these two types of men ovide some clues to imaging studies of encoding and retrieval in LTM.for to Brodmann areas 44. 5 and 47.DLFC to areas 9 and 4 and 10.It is our inter likelyto entail maintaining and manipulating ever, on in WN ded into LTM. pic sulcal/gyral features (ont which th that similar FC dissociat ons of function have been PET and fMRI activations are mapped)and the bounda both LTM and WM imaging studies, yet these (Rolan et eptions (Wagner 1999) diffe in the size of Broca's are oduce the evidence is to consider each field sep 10 individuals,the microscopic boundaries bearing litt before subsequently discussing how they may converge stent relationship to mac scopic landmarks (Amunts We therefore address the patterns of memory-related F et al,1999 exerc n re Pirst,we Brodmann's definitions.The chosen subdivisions are also LTM retrieval).Secondly.in the likely to reflect differences in patterns of connectivity oncluding section.we attempt a more general interpretation (Passingham.1993:Fuster. 1997).0nce more,however. that extends to FC activations across the different domains Frontal lobes and human memory 851 Fig. 1 Left-sided view of human brain showing our working subdivisions of lateral FC. The border between VLFC and DLFC is marked by the inferior frontal sulcus. The posterior border of AFC is marked by a line drawn vertically at the anterior edge of the inferior frontal gyrus. distinctions (in addition to the left–right lateralization of the we cannot be certain of the precise relationship between regions) with due consideration of the imperfect spatial connectivity and macroanatomical landmarks, and we refrain resolution of the techniques, of the enormous anatomical from further speculation. Finally, we confine our review to variability among subjects, and of the likelihood that, studies of groups of young, healthy individuals. ultimately, these broad areas will themselves be shown to be The nature of the contribution of the frontal lobe to functionally subdivided. The rationale behind this division memory is clouded by the division of the experimental is, on the one hand, an attempt to acknowledge the limited literature into two broad fields: working memory (WM), the spatial information provided by group studies (particularly ability to maintain information temporarily over periods of with PET) and, on the other hand, to avoid treating clearly seconds, and long-term memory (LTM), the ability to retain separate regional responses as undifferentiated ‘frontal’ information for much longer periods. While there are good activations. Our particular subdivisions are based on existing reasons for distinguishing between these two types of memfunctional imaging data rather than microstructural findings, ory, it is also likely that considerable overlap exists between although they may be considered to provide some clues to the frontally mediated processes involved in each. Many the underlying anatomy. Thus, VLFC corresponds loosely imaging studies of encoding and retrieval in LTM, for to Brodmann areas 44, 45 and 47, DLFC to areas 9 and 46 example, are likely to entail maintaining and manipulating and AFC to areas 8 and 10. It is our intention, however, to information in WM. Conversely, information maintained in avoid relying upon the uncertain and inconsistent relationship WM may be encoded into LTM. It is interesting, therefore, between macroscopic sulcal/gyral features (onto which the that similar FC dissociations of function have been proposed PET and fMRI activations are mapped) and the boundaries in both LTM and WM imaging studies, and yet these of the Brodmann areas (Roland et al., 1997; Zilles et al., findings, with certain exceptions (Wagner, 1999), are not 1997). Amunts and colleagues, for example, noted a 10-fold often considered together. Nonetheless, a convenient way to difference in the size of Broca’s area across a group of introduce the evidence is to consider each field separately, 10 individuals, the microscopic boundaries bearing little before subsequently discussing how they may converge. consistent relationship to macroscopic landmarks (Amunts We therefore address the patterns of memory-related FC et al., 1999). Caution must therefore be exercised in relating activation in two stages. First, we consider interpretations of macro- to microanatomy, and we will avoid the use of FC activations offered by researchers within each domain Brodmann’s definitions. The chosen subdivisions are also (WM, LTM encoding, LTM retrieval). Secondly, in the likely to reflect differences in patterns of connectivity concluding section, we attempt a more general interpretation (Passingham, 1993; Fuster, 1997). Once more, however, that extends to FC activations across the different domains