正在加载图片...
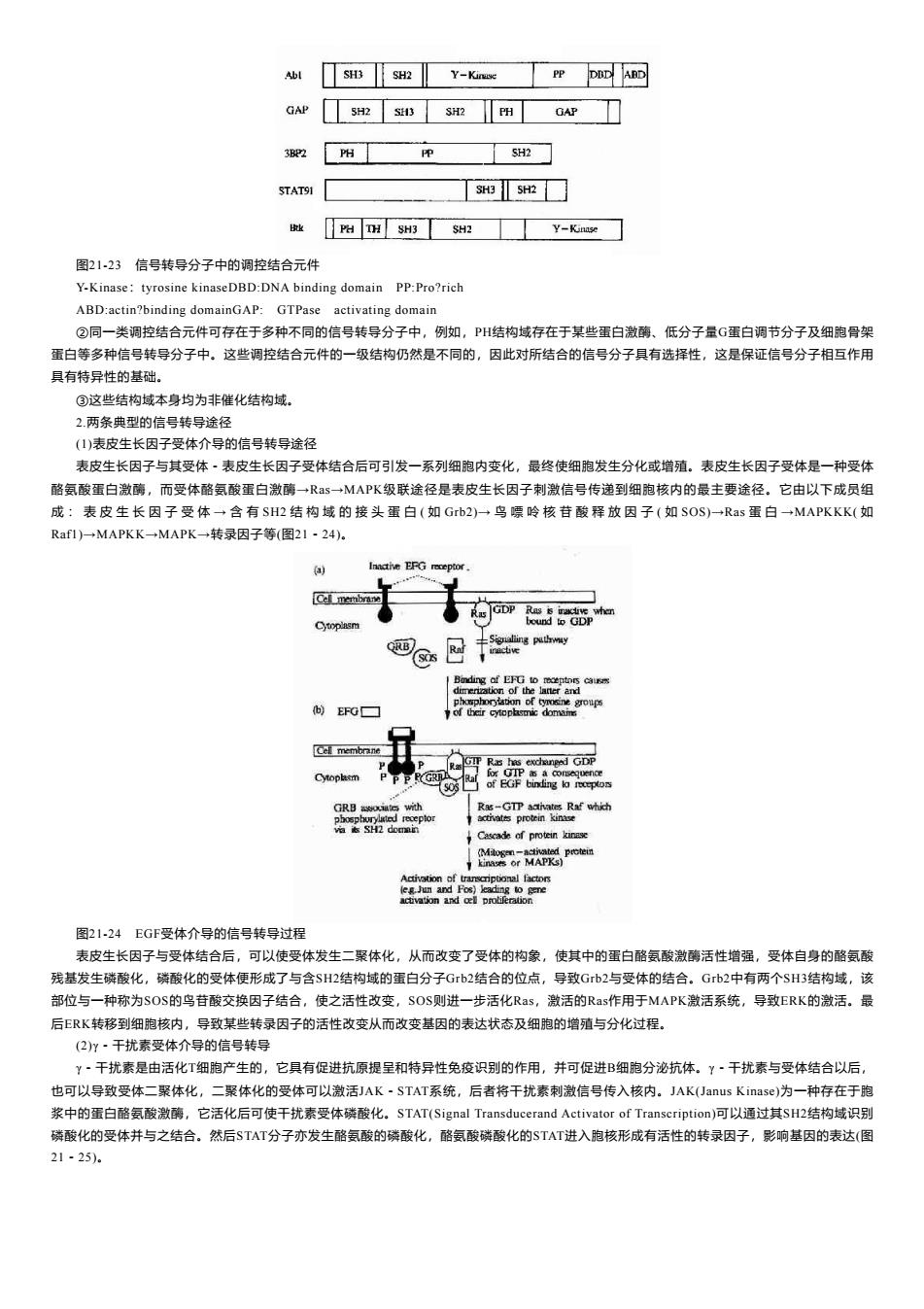
Abl SH3 SH2 Y-Kinase PP DDD ARD GAP SH2 S3 SH2 阳 GAP 3B2 H P SH2 STAT91 SH3 SH2 PH TH SH3 SH2 Y-Kinase 图21-23信号转导分子中的调控结合元件 Y-Kinase:tyrosine kinaseDBD:DNA binding domain PP:Pro?rich ABD:actin?binding domainGAP:GTPase activating domain ②同一类调控结合元件可存在于多种不同的信号转导分子中,例如,PH结构域存在于某些蛋白激酶、低分子量G蛋白调节分子及细胞骨架 蛋白等多种信号转导分子中。这些调控结合元件的一级结构仍然是不同的,因此对所结合的信号分子具有选择性,这是保证信号分子相互作用 具有特异性的基础。 ③这些结构域本身均为非催化结构域。 2两条典型的信号转导途径 (1)表皮生长因子受体介导的信号转导途径 表皮生长因子与其受体·表皮生长因子受体结合后可引发一系列细胞内变化,最终使细胞发生分化或增殖。表皮生长因子受体是一种受体 酪氨酸蛋白激酶,而受体酪氨酸蛋白激酶一Rs一MAPK级联途径是表皮生长因子刺激信号传递到细胞核内的最主要途径。它由以下成员组 成:表皮生长因子受体一含有SH2结构域的接头蛋白(如Grb2)一鸟嘌呤核苷酸释放因子(如SOS)→Rs蛋白一MAPKKK(如 Raf1)→MAPKK→MAPK→转录因子等(图21·24). Inactive EFG receptor GDP 主Signaling puthwy 向)EFG☐ Cell membrane GRB aoi琪owh Ras-GTP activates Raf which phosphurylted receptor activates protein kinase via is SH2 domin Cascade of protein kinase (Mogen-activated prctein kinases or MAPKs) Activation of transcriptional factors leg.Jun and Fos)leading to gene activation and oel protiferation 图21-24EGF受体介导的信号转导过程 表皮生长因子与受体结合后,可以使受体发生二聚体化,从而改变了受体的构象,使其中的蛋白酪氨酸激酶活性增强,受体自身的酪氨酸 残基发生磷酸化,磷酸化的受体便形成了与含SH2结构域的蛋白分子Gb2结合的位点,导致Gb2与受体的结合。Gb2中有两个SH3结构域,该 部位与一种称为SOS的鸟苷酸交换因子结合,使之活性改变,SOS则进一步活化Ras,激活的Ras作用于MAPK激活系统,导致ERK的激活。最 后ERK转移到细胞核内,导致某些转录因子的活性改变从而改变基因的表达状态及细胞的增殖与分化过程。 (2)y·干扰素受体介导的信号转导 Y·干扰素是由活化T细胞产生的,它具有促进抗原提呈和特异性免疫识别的作用,并可促进B细胞分泌抗体。Y·干扰素与受体结合以后, 也可以导致受体二聚体化,二聚体化的受体可以激活JAK·STAT系统,后者将干扰素刺激信号传入核内。JAK(Janus Kinase))为一种存在于胞 浆中的蛋白酪氨酸激酶,它活化后可使干扰素受体磷酸化。STAT(Signal Transducerand Activator of Transcription)可以通过其SH2结构域识别 磷酸化的受体并与之结合。然后$TAT分子亦发生酪氨酸的磷酸化,酪氨酸磷酸化的$TAT进入胞核形成有话性的转录因子,影响基因的表达(图 21-25).图21-23 信号转导分子中的调控结合元件 Y-Kinase:tyrosine kinaseDBD:DNA binding domain PP:Pro?rich ABD:actin?binding domainGAP: GTPase activating domain ②同一类调控结合元件可存在于多种不同的信号转导分子中,例如,PH结构域存在于某些蛋白激酶、低分子量G蛋白调节分子及细胞骨架 蛋白等多种信号转导分子中。这些调控结合元件的一级结构仍然是不同的,因此对所结合的信号分子具有选择性,这是保证信号分子相互作用 具有特异性的基础。 ③这些结构域本身均为非催化结构域。 2.两条典型的信号转导途径 (1)表皮生长因子受体介导的信号转导途径 表皮生长因子与其受体-表皮生长因子受体结合后可引发一系列细胞内变化,最终使细胞发生分化或增殖。表皮生长因子受体是一种受体 酪氨酸蛋白激酶,而受体酪氨酸蛋白激酶→Ras→MAPK级联途径是表皮生长因子刺激信号传递到细胞核内的最主要途径。它由以下成员组 成 : 表 皮 生 长 因 子 受 体 → 含 有 SH2 结 构 域 的 接 头 蛋 白 ( 如 Grb2)→ 鸟 嘌 呤 核 苷 酸 释 放 因 子 ( 如 SOS)→Ras 蛋 白 →MAPKKK( 如 Raf1)→MAPKK→MAPK→转录因子等(图21-24)。 图21-24 EGF受体介导的信号转导过程 表皮生长因子与受体结合后,可以使受体发生二聚体化,从而改变了受体的构象,使其中的蛋白酪氨酸激酶活性增强,受体自身的酪氨酸 残基发生磷酸化,磷酸化的受体便形成了与含SH2结构域的蛋白分子Grb2结合的位点,导致Grb2与受体的结合。Grb2中有两个SH3结构域,该 部位与一种称为SOS的鸟苷酸交换因子结合,使之活性改变,SOS则进一步活化Ras,激活的Ras作用于MAPK激活系统,导致ERK的激活。最 后ERK转移到细胞核内,导致某些转录因子的活性改变从而改变基因的表达状态及细胞的增殖与分化过程。 (2)γ-干扰素受体介导的信号转导 γ-干扰素是由活化T细胞产生的,它具有促进抗原提呈和特异性免疫识别的作用,并可促进B细胞分泌抗体。γ-干扰素与受体结合以后, 也可以导致受体二聚体化,二聚体化的受体可以激活JAK-STAT系统,后者将干扰素刺激信号传入核内。JAK(Janus Kinase)为一种存在于胞 浆中的蛋白酪氨酸激酶,它活化后可使干扰素受体磷酸化。STAT(Signal Transducerand Activator of Transcription)可以通过其SH2结构域识别 磷酸化的受体并与之结合。然后STAT分子亦发生酪氨酸的磷酸化,酪氨酸磷酸化的STAT进入胞核形成有活性的转录因子,影响基因的表达(图 21-25)