正在加载图片...
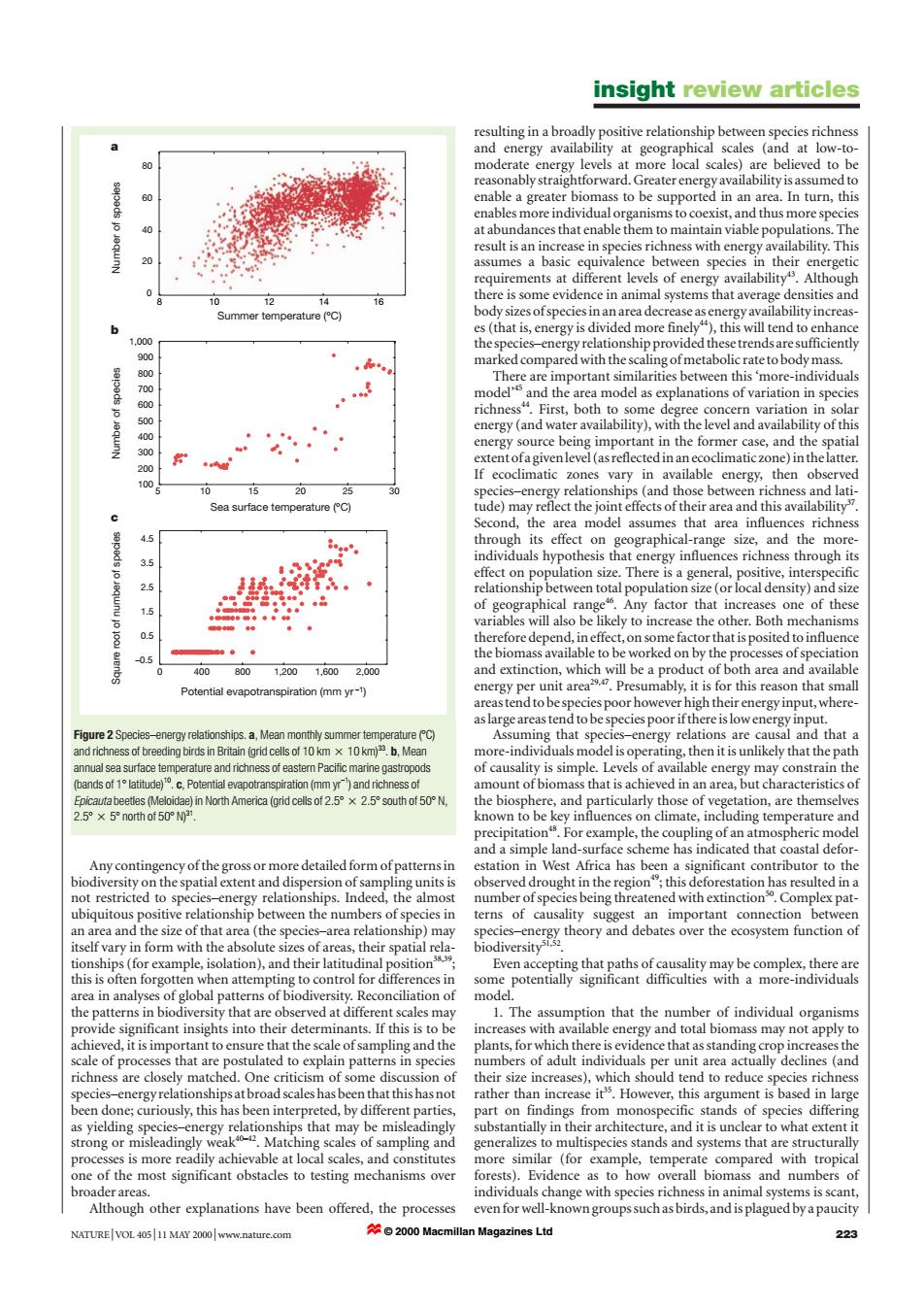
insight review articles resulting in a broadly positive relationship between species richness and energy availability at geographical scales (and at low-to- moderate energy levels at more local scales)are believed to be reasonably straightforward.Greater energy availability is assumed to enable a greater biomass to be supported in an area.In turn,this enables more individual organisms to coexist,and thus more species at abundances that enable them to maintain viable populations.The result is an increase in species richness with energy availability.This assumes a basic equivalence between species in their energetic requirements at different levels of energy availability.Although 12 14 there is some evidence in animal systems that average densities and Summer temperature(C) body sizes ofspecies in an area decrease as energy availability increas- es(that is,energy is divided more finely4),this will tend to enhance 000 the species-energy relationship provided these trends are sufficiently 900 marked compared with the scaling of metabolic rate to body mass. 800 There are important similarities between this 'more-individuals 700 model'and the area model as explanations of variation in species 600 richness".First,both to some degree concern variation in solar energy(and water availability),with the level and availability of this 300 energy source being important in the former case,and the spatial extent ofa givenlevel(as reflected in an ecoclimatic zone)in thelatter. 200 100 If ecoclimatic zones vary in available energy,then observed 10 15 20 species-energy relationships(and those between richness and lati- Sea surface temperature (C) tude)may reflect the joint effects of their area and this availability Second,the area model assumes that area influences richness through its effect on geographical-range size,and the more- 35 individuals hypothesis that energy influences richness through its effect on population size.There is a general,positive,interspecific 25 relationship between total population size(or local density)and size of geographical range.Any factor that increases one of these 5 variables will also be likely to increase the other.Both mechanisms 05 therefore depend,in effect,on some factor that is posited to influence the biomass available to be worked on by the processes of speciation 0.5 400 8001.2001.6002.000 and extinction,which will be a product of both area and available Potential evapotranspiration(mm yr-1) energy per unit area Presumably,it is for this reason that small areas tend to be species poor however high their energy input,where- as large areas tend to be species poor if there is low energy input. Figure 2 Species-energy relationships.a,Mean monthly summer temperature(C) Assuming that species-energy relations are causal and that a and richness of breeding birds in Britain (grid cells of 10 km x 10 km).b,Mean more-individuals model is operating,then it is unlikely that the path annual sea surface temperature and richness of eastern Pacific marine gastropods of causality is simple.Levels of available energy may constrain the (bands of 1 latitude).c.Potential evapotranspiration (mm yr)and richness of amount of biomass that is achieved in an area,but characteristics of Epicautabeetles (Meloidae)in North America(grid cells of2.5°×2.5°south of50°N, the biosphere,and particularly those of vegetation,are themselves 2.5°×5°n0thof50°031 known to be key influences on climate,including temperature and precipitation.For example,the coupling of an atmospheric model and a simple land-surface scheme has indicated that coastal defor- Any contingency ofthe gross or more detailed form of patterns in estation in West Africa has been a significant contributor to the biodiversity on the spatial extent and dispersion of sampling units is observed drought in the region;this deforestation has resulted in a not restricted to species-energy relationships.Indeed,the almost number of species being threatened with extinction.Complex pat- ubiquitous positive relationship between the numbers of species in terns of causality suggest an important connection between an area and the size of that area (the species-area relationship)may species-energy theory and debates over the ecosystem function of itself vary in form with the absolute sizes of areas,their spatial rela- biodiversity1.52 tionships(for example,isolation),and their latitudinal position Even accepting that paths of causality may be complex,there are this is often forgotten when attempting to control for differences in some potentially significant difficulties with a more-individuals area in analyses of global patterns of biodiversity.Reconciliation of model. the patterns in biodiversity that are observed at different scales may 1.The assumption that the number of individual organisms provide significant insights into their determinants.If this is to be increases with available energy and total biomass may not apply to achieved,it is important to ensure that the scale of sampling and the plants,for which there is evidence that as standing crop increases the scale of processes that are postulated to explain patterns in species numbers of adult individuals per unit area actually declines (and richness are closely matched.One criticism of some discussion of their size increases),which should tend to reduce species richness species-energy relationships at broad scales has been that this has not rather than increase it.However,this argument is based in large been done;curiously,this has been interpreted,by different parties, part on findings from monospecific stands of species differing as yielding species-energy relationships that may be misleadingly substantially in their architecture,and it is unclear to what extent it strong or misleadingly weak.Matching scales of sampling and generalizes to multispecies stands and systems that are structurally processes is more readily achievable at local scales,and constitutes more similar (for example,temperate compared with tropical one of the most significant obstacles to testing mechanisms over forests).Evidence as to how overall biomass and numbers of broader areas. individuals change with species richness in animal systems is scant, Although other explanations have been offered,the processes even for well-known groupssuchas birds,and is plagued by apaucity NATURE|VOL 40511 MAY 2000 www.nature.com ☆©20o0 Macmillan Magazines Ltd 223Any contingency of the gross or more detailed form of patterns in biodiversity on the spatial extent and dispersion of sampling units is not restricted to species–energy relationships. Indeed, the almost ubiquitous positive relationship between the numbers of species in an area and the size of that area (the species–area relationship) may itself vary in form with the absolute sizes of areas, their spatial relationships (for example, isolation), and their latitudinal position38,39; this is often forgotten when attempting to control for differences in area in analyses of global patterns of biodiversity. Reconciliation of the patterns in biodiversity that are observed at different scales may provide significant insights into their determinants. If this is to be achieved, it is important to ensure that the scale of sampling and the scale of processes that are postulated to explain patterns in species richness are closely matched. One criticism of some discussion of species–energy relationships at broad scales has been that this has not been done; curiously, this has been interpreted, by different parties, as yielding species–energy relationships that may be misleadingly strong or misleadingly weak40–42. Matching scales of sampling and processes is more readily achievable at local scales, and constitutes one of the most significant obstacles to testing mechanisms over broader areas. Although other explanations have been offered, the processes resulting in a broadly positive relationship between species richness and energy availability at geographical scales (and at low-tomoderate energy levels at more local scales) are believed to be reasonably straightforward. Greater energy availability is assumed to enable a greater biomass to be supported in an area. In turn, this enables more individual organisms to coexist, and thus more species at abundances that enable them to maintain viable populations. The result is an increase in species richness with energy availability. This assumes a basic equivalence between species in their energetic requirements at different levels of energy availability43. Although there is some evidence in animal systems that average densities and body sizes of species in an area decrease as energy availability increases (that is, energy is divided more finely44), this will tend to enhance the species–energy relationship provided these trends are sufficiently marked compared with the scaling of metabolic rate to body mass. There are important similarities between this ‘more-individuals model’45 and the area model as explanations of variation in species richness44. First, both to some degree concern variation in solar energy (and water availability), with the level and availability of this energy source being important in the former case, and the spatial extent of a given level (as reflected in an ecoclimatic zone) in the latter. If ecoclimatic zones vary in available energy, then observed species–energy relationships (and those between richness and latitude) may reflect the joint effects of their area and this availability37. Second, the area model assumes that area influences richness through its effect on geographical-range size, and the moreindividuals hypothesis that energy influences richness through its effect on population size. There is a general, positive, interspecific relationship between total population size (or local density) and size of geographical range46. Any factor that increases one of these variables will also be likely to increase the other. Both mechanisms therefore depend, in effect, on some factor that is posited to influence the biomass available to be worked on by the processes of speciation and extinction, which will be a product of both area and available energy per unit area29,47. Presumably, it is for this reason that small areas tend to be species poor however high their energy input, whereas large areas tend to be species poor if there is low energy input. Assuming that species–energy relations are causal and that a more-individuals model is operating, then it is unlikely that the path of causality is simple. Levels of available energy may constrain the amount of biomass that is achieved in an area, but characteristics of the biosphere, and particularly those of vegetation, are themselves known to be key influences on climate, including temperature and precipitation48. For example, the coupling of an atmospheric model and a simple land-surface scheme has indicated that coastal deforestation in West Africa has been a significant contributor to the observed drought in the region49; this deforestation has resulted in a number of species being threatened with extinction50. Complex patterns of causality suggest an important connection between species–energy theory and debates over the ecosystem function of biodiversity51,52. Even accepting that paths of causality may be complex, there are some potentially significant difficulties with a more-individuals model. 1. The assumption that the number of individual organisms increases with available energy and total biomass may not apply to plants, for which there is evidence that as standing crop increases the numbers of adult individuals per unit area actually declines (and their size increases), which should tend to reduce species richness rather than increase it35. However, this argument is based in large part on findings from monospecific stands of species differing substantially in their architecture, and it is unclear to what extent it generalizes to multispecies stands and systems that are structurally more similar (for example, temperate compared with tropical forests). Evidence as to how overall biomass and numbers of individuals change with species richness in animal systems is scant, even for well-known groups such as birds, and is plagued by a paucity insight review articles NATURE | VOL 405 | 11 MAY 2000 | www.nature.com 223 Summer temperature (°C) Number of species Number of species 0 20 40 60 80 8 10 12 14 16 Sea surface temperature (°C) Potential evapotranspiration (mm yr –1) 100 200 300 400 500 600 700 800 900 1,000 5 10 15 20 25 30 Square root of number of species –0.5 0.5 1.5 2.5 3.5 4.5 0 400 800 1,200 1,600 2,000 a b c Figure 2 Species–energy relationships. a, Mean monthly summer temperature (7C) and richness of breeding birds in Britain (grid cells of 10 km 2 10 km)33. b, Mean annual sea surface temperature and richness of eastern Pacific marine gastropods (bands of 17 latitude)10. c, Potential evapotranspiration (mm yr–1) and richness of Epicauta beetles (Meloidae) in North America (grid cells of 2.57 2 2.57south of 507 N, 2.57 2 57 north of 507 N)31. © 2000 Macmillan Magazines Ltd