正在加载图片...
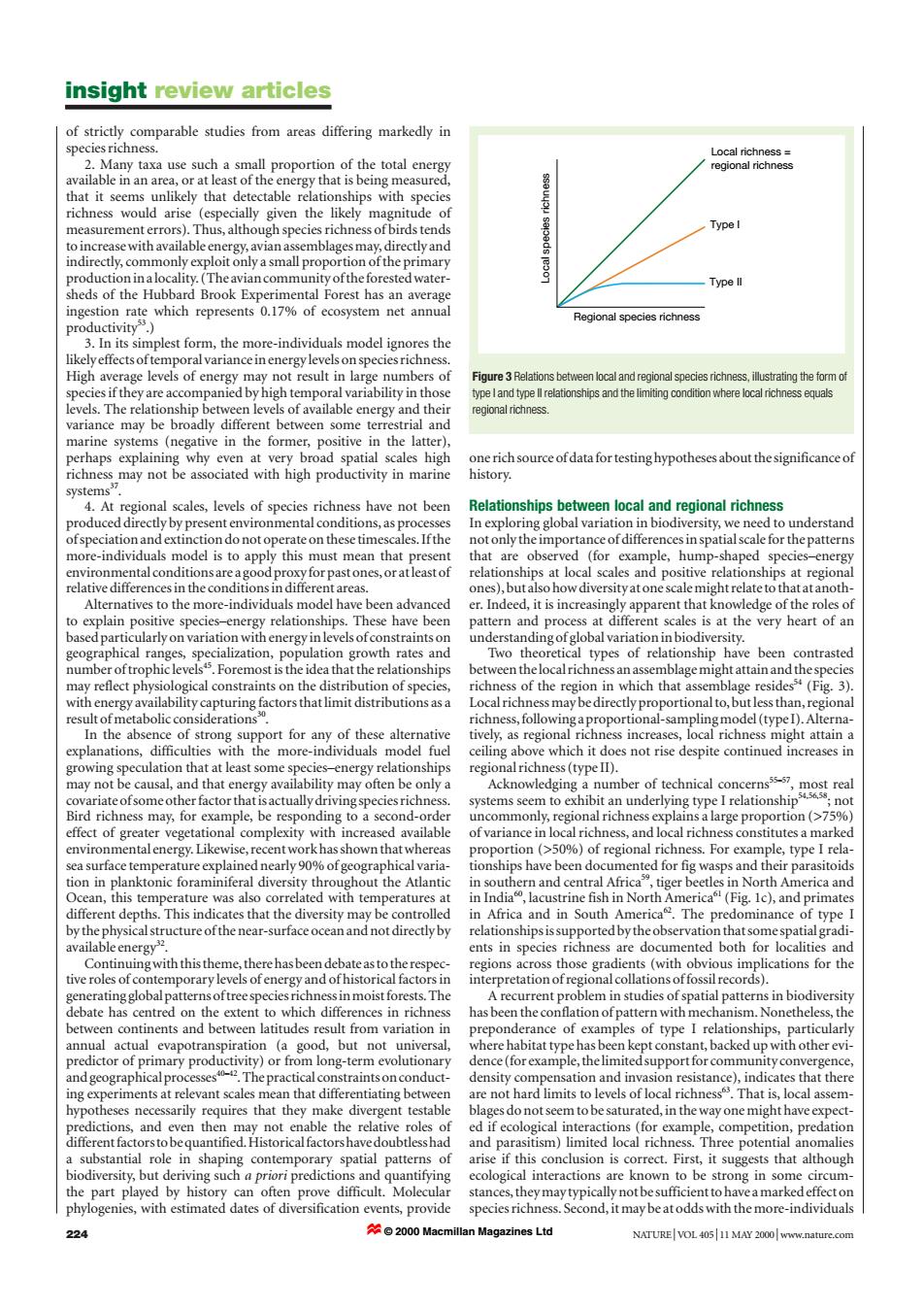
insight review articles of strictly comparable studies from areas differing markedly in species richness. Local richness 2.Many taxa use such a small proportion of the total energy regional richness available in an area,or at least of the energy that is being measured, that it seems unlikely that detectable relationships with species richness would arise (especially given the likely magnitude of measurement errors).Thus,although species richness of birds tends Type I to increase with available energy,avian assemblages may,directly and indirectly,commonly exploit only a small proportion of the primary production ina locality.(The avian community ofthe forested water- Type ll sheds of the Hubbard Brook Experimental Forest has an average ingestion rate which represents 0.17%of ecosystem net annual Regional species richness productivity) 3.In its simplest form,the more-individuals model ignores the likely effects oftemporal variance in energy levels on species richness. High average levels of energy may not result in large numbers of Figure 3 Relations between local and regional species richness,illustrating the form of species if they are accompanied by high temporal variability in those type Iand type ll relationships and the limiting condition where local richness equals levels.The relationship between levels of available energy and their regional richness. variance may be broadly different between some terrestrial and marine systems (negative in the former,positive in the latter), perhaps explaining why even at very broad spatial scales high one rich source ofdata for testing hypotheses about the significance of richness may not be associated with high productivity in marine history. systems37 4.At regional scales,levels of species richness have not been Relationships between local and regional richness produced directly by present environmental conditions,as processes In exploring global variation in biodiversity,we need to understand ofspeciation and extinction do not operate on these timescales.Ifthe not only the importance of differences in spatial scale for the patterns more-individuals model is to apply this must mean that present that are observed (for example,hump-shaped species-energy environmental conditionsare a good proxy for past ones,or atleast of relationships at local scales and positive relationships at regional relative differences in the conditions in different areas. ones),but also how diversity at one scale might relate to that at anoth- Alternatives to the more-individuals model have been advanced er.Indeed,it is increasingly apparent that knowledge of the roles of to explain positive species-energy relationships.These have been pattern and process at different scales is at the very heart of an based particularly on variation with energy in levels of constraints on understanding ofglobal variation in biodiversity. geographical ranges,specialization,population growth rates and Two theoretical types of relationship have been contrasted number oftrophiclevels5.Foremost is the idea that the relationships between thelocal richness an assemblage might attain and the species may reflect physiological constraints on the distribution of species, richness of the region in which that assemblage residess(Fig.3). with energy availability capturing factors that limit distributions as a Local richness may bedirectly proportional to,but less than,regional result of metabolic considerations. richness,followingaproportional-samplingmodel(typeI).Alterna- In the absence of strong support for any of these alternative tively,as regional richness increases,local richness might attain a explanations,difficulties with the more-individuals model fuel ceiling above which it does not rise despite continued increases in growing speculation that at least some species-energy relationships regional richness(type II). may not be causal,and that energy availability may often be only a Acknowledging a number of technical concerns35-57,most real covariate ofsome other factor that is actually driving species richness. systems seem to exhibit an underlying type I relationship ;not Bird richness may,for example,be responding to a second-order uncommonly,regional richness explains alarge proportion(>75%) effect of greater vegetational complexity with increased available of variance in local richness,and local richness constitutes a marked environmental energy.Likewise,recent workhas shown that whereas proportion (>50%)of regional richness.For example,type I rela- sea surface temperature explained nearly 90%of geographical varia- tionships have been documented for fig wasps and their parasitoids tion in planktonic foraminiferal diversity throughout the Atlantic in southern and central Africa",tiger beetles in North America and Ocean,this temperature was also correlated with temperatures at in India,lacustrine fish in North America(Fig.Ic),and primates different depths.This indicates that the diversity may be controlled in Africa and in South America.The predominance of type I by the physical structure ofthe near-surface ocean and not directly by relationships is supported by the observation that some spatial gradi- available energy2. ents in species richness are documented both for localities and Continuing with this theme,there has been debate as to the respec- regions across those gradients (with obvious implications for the tive roles of contemporary levels ofenergy and ofhistorical factors in interpretation of regional collations offossil records). generatingglobalpatternsoftreespeciesrichnessinmoist forests.The A recurrent problem in studies of spatial patterns in biodiversity debate has centred on the extent to which differences in richness has been the conflation of pattern with mechanism.Nonetheless,the between continents and between latitudes result from variation in preponderance of examples of type I relationships,particularly annual actual evapotranspiration (a good,but not universal, where habitat type has been kept constant,backed up with other evi- predictor of primary productivity)or from long-term evolutionary dence(forexample,thelimitedsupport for communityconvergence, and geographicalprocesses-.The practical constraintsonconduct- density compensation and invasion resistance),indicates that there ing experiments at relevant scales mean that differentiating between are not hard limits to levels of local richness.That is,local assem- hypotheses necessarily requires that they make divergent testable blages do not seem to be saturated,in the way one might have expect- predictions,and even then may not enable the relative roles of ed if ecological interactions (for example,competition,predation different factors to be quantified.Historical factors have doubtless had and parasitism)limited local richness.Three potential anomalies a substantial role in shaping contemporary spatial patterns of arise if this conclusion is correct.First,it suggests that although biodiversity,but deriving such a priori predictions and quantifying ecological interactions are known to be strong in some circum- the part played by history can often prove difficult.Molecular stances,they may typically not be sufficient to have a marked effect on phylogenies,with estimated dates of diversification events,provide species richness.Second,it may be at odds with the more-individuals 224 2000 Macmillan Magazines Ltd NATURE VOL 40511 MAY 2000 www.nature.comof strictly comparable studies from areas differing markedly in species richness. 2. Many taxa use such a small proportion of the total energy available in an area, or at least of the energy that is being measured, that it seems unlikely that detectable relationships with species richness would arise (especially given the likely magnitude of measurement errors). Thus, although species richness of birds tends to increase with available energy, avian assemblages may, directly and indirectly, commonly exploit only a small proportion of the primary production in a locality. (The avian community of the forested watersheds of the Hubbard Brook Experimental Forest has an average ingestion rate which represents 0.17% of ecosystem net annual productivity53.) 3. In its simplest form, the more-individuals model ignores the likely effects of temporal variance in energy levels on species richness. High average levels of energy may not result in large numbers of species if they are accompanied by high temporal variability in those levels. The relationship between levels of available energy and their variance may be broadly different between some terrestrial and marine systems (negative in the former, positive in the latter), perhaps explaining why even at very broad spatial scales high richness may not be associated with high productivity in marine systems37. 4. At regional scales, levels of species richness have not been produced directly by present environmental conditions, as processes of speciation and extinction do not operate on these timescales. If the more-individuals model is to apply this must mean that present environmental conditions are a good proxy for past ones, or at least of relative differences in the conditions in different areas. Alternatives to the more-individuals model have been advanced to explain positive species–energy relationships. These have been based particularly on variation with energy in levels of constraints on geographical ranges, specialization, population growth rates and number of trophic levels45. Foremost is the idea that the relationships may reflect physiological constraints on the distribution of species, with energy availability capturing factors that limit distributions as a result of metabolic considerations30. In the absence of strong support for any of these alternative explanations, difficulties with the more-individuals model fuel growing speculation that at least some species–energy relationships may not be causal, and that energy availability may often be only a covariate of some other factor that is actually driving species richness. Bird richness may, for example, be responding to a second-order effect of greater vegetational complexity with increased available environmental energy. Likewise, recent work has shown that whereas sea surface temperature explained nearly 90% of geographical variation in planktonic foraminiferal diversity throughout the Atlantic Ocean, this temperature was also correlated with temperatures at different depths. This indicates that the diversity may be controlled by the physical structure of the near-surface ocean and not directly by available energy32. Continuing with this theme, there has been debate as to the respective roles of contemporary levels of energy and of historical factors in generating global patterns of tree species richness in moist forests. The debate has centred on the extent to which differences in richness between continents and between latitudes result from variation in annual actual evapotranspiration (a good, but not universal, predictor of primary productivity) or from long-term evolutionary and geographical processes40–42. The practical constraints on conducting experiments at relevant scales mean that differentiating between hypotheses necessarily requires that they make divergent testable predictions, and even then may not enable the relative roles of different factors to be quantified. Historical factors have doubtless had a substantial role in shaping contemporary spatial patterns of biodiversity, but deriving such a priori predictions and quantifying the part played by history can often prove difficult. Molecular phylogenies, with estimated dates of diversification events, provide one rich source of data for testing hypotheses about the significance of history. Relationships between local and regional richness In exploring global variation in biodiversity, we need to understand not only the importance of differences in spatial scale for the patterns that are observed (for example, hump-shaped species–energy relationships at local scales and positive relationships at regional ones), but also how diversity at one scale might relate to that at another. Indeed, it is increasingly apparent that knowledge of the roles of pattern and process at different scales is at the very heart of an understanding of global variation in biodiversity. Two theoretical types of relationship have been contrasted between the local richness an assemblage might attain and the species richness of the region in which that assemblage resides54 (Fig. 3). Local richness may be directly proportional to, but less than, regional richness, following a proportional-sampling model (type I). Alternatively, as regional richness increases, local richness might attain a ceiling above which it does not rise despite continued increases in regional richness (type II). Acknowledging a number of technical concerns55–57, most real systems seem to exhibit an underlying type I relationship54,56,58; not uncommonly, regional richness explains a large proportion (>75%) of variance in local richness, and local richness constitutes a marked proportion (>50%) of regional richness. For example, type I relationships have been documented for fig wasps and their parasitoids in southern and central Africa59, tiger beetles in North America and in India60, lacustrine fish in North America61 (Fig. 1c), and primates in Africa and in South America62. The predominance of type I relationships is supported by the observation that some spatial gradients in species richness are documented both for localities and regions across those gradients (with obvious implications for the interpretation of regional collations of fossil records). A recurrent problem in studies of spatial patterns in biodiversity has been the conflation of pattern with mechanism. Nonetheless, the preponderance of examples of type I relationships, particularly where habitat type has been kept constant, backed up with other evidence (for example, the limited support for community convergence, density compensation and invasion resistance), indicates that there are not hard limits to levels of local richness63. That is, local assemblages do not seem to be saturated, in the way one might have expected if ecological interactions (for example, competition, predation and parasitism) limited local richness. Three potential anomalies arise if this conclusion is correct. First, it suggests that although ecological interactions are known to be strong in some circumstances, they may typically not be sufficient to have a marked effect on species richness. Second, it may be at odds with the more-individuals insight review articles 224 NATURE | VOL 405 | 11 MAY 2000 | www.nature.com Regional species richness Local species richness Local richness = regional richness Type I Type II Figure 3 Relations between local and regional species richness, illustrating the form of type I and type II relationships and the limiting condition where local richness equals regional richness. © 2000 Macmillan Magazines Ltd