正在加载图片...
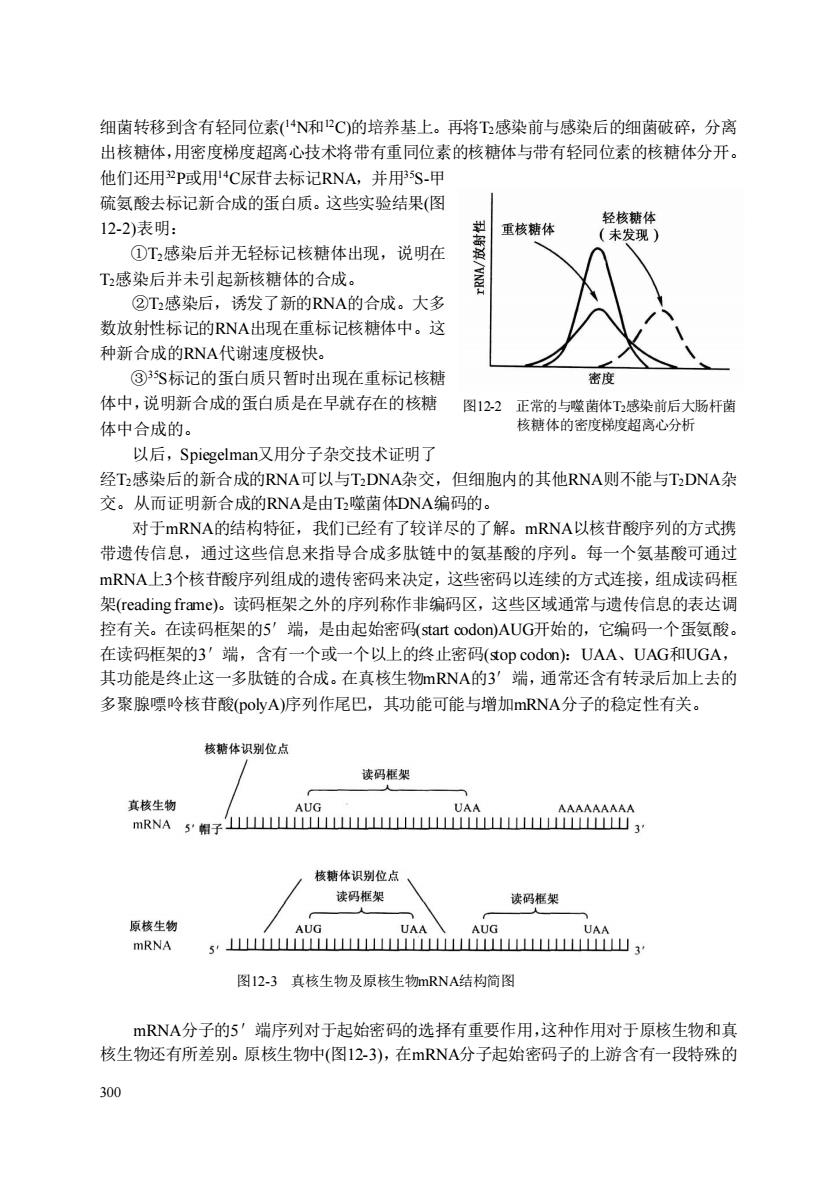
细菌转移到含有轻同位素(N和P℃)的培养基上。再将T感染前与感染后的细菌破碎,分离 出核糖体,用密度梯度超离心技术将带有重同位素的核糖体与带有轻同位素的核糖体分开。 他们还用2P或用4C尿苷去标记RNA,并用5S-甲 硫氨酸去标记新合成的蛋白质。这些实验结果(图 12-2)表明: 重核糖体 ①T2感染后并无轻标记核糖体出现,说明在 T2感染后并未引起新核糖体的合成。 ②T感染后,诱发了新的RNA的合成。大多 数放射性标记的RNA出现在重标记核糖体中。这 种新合成的RNA代谢速度极快。 ③S标记的蛋白质只暂时出现在重标记核糖 密度 体中,说明新合成的蛋白质是在早就存在的核糖图12.2正常的与噬菌体T感染前后大肠杆菌 体中合成的。 核糖体的密度梯度超离心分析 以后,Spiegelman又用分子杂交技术证明了 经T感染后的新合成的RNA可以与TDNA杂交,但细胞内的其他RNA则不能与TDNA杂 交。从而证明新合成的RNA是由T噬菌体DNA编码的。 对于mRNA的结构特征,我们已经有了较详尽的了解。mRNA以核苷酸序列的方式携 带遗传信息,通过这些信息来指导合成多肽链中的氨基酸的序列。每一个氨基酸可通过 mRNA上3个核苷酸序列组成的遗传密码来决定,这些密码以连续的方式连接,组成读码框 架(reading frame)。读码框架之外的序列称作非编码区,这些区域通常与遗传信息的表达调 控有关。在读码框架的5'端,是由起始密码((start codon)AUG开始的,它编码一个蛋氨酸 在读码框架的3'端,含有一个或一个以上的终止密码(top codon:UAA、UAG和UGA, 其功能是终止这一多肽链的合成。在真核生物mRNA的3'端,通常还含有转录后加上去的 多聚腺票岭核苷酸(Do小vA)序列作尾巴,其功能可能与增加mRNA分子的隐定性有关。 核糖体识别位店 读码框架 真核生物 mRNA 5帽子山YMM 核糖体识别位点 读码框架 读码据架 原核生物 mRNA s'出i山, 图12-3真核生物及原核生物mRNA结构简图 mRNA分子的5'端序列对于起始密码的选择有重要作用,这种作用对于原核生物和真 核生物还有所差别。原核生物中(图12-3),在mRNA分子起始密码子的上游含有一段特殊的 300 300 细菌转移到含有轻同位素( 14N和12C)的培养基上。再将T2感染前与感染后的细菌破碎,分离 出核糖体,用密度梯度超离心技术将带有重同位素的核糖体与带有轻同位素的核糖体分开。 他们还用32P或用14C尿苷去标记RNA,并用35S-甲 硫氨酸去标记新合成的蛋白质。这些实验结果(图 12-2)表明: ①T2感染后并无轻标记核糖体出现,说明在 T2感染后并未引起新核糖体的合成。 ②T2感染后,诱发了新的RNA的合成。大多 数放射性标记的RNA出现在重标记核糖体中。这 种新合成的RNA代谢速度极快。 ③35S标记的蛋白质只暂时出现在重标记核糖 体中,说明新合成的蛋白质是在早就存在的核糖 体中合成的。 以后,Spiegelman又用分子杂交技术证明了 经T2感染后的新合成的RNA可以与T2DNA杂交,但细胞内的其他RNA则不能与T2DNA杂 交。从而证明新合成的RNA是由T2噬菌体DNA编码的。 对于mRNA的结构特征,我们已经有了较详尽的了解。mRNA以核苷酸序列的方式携 带遗传信息,通过这些信息来指导合成多肽链中的氨基酸的序列。每一个氨基酸可通过 mRNA上3个核苷酸序列组成的遗传密码来决定,这些密码以连续的方式连接,组成读码框 架(reading frame)。读码框架之外的序列称作非编码区,这些区域通常与遗传信息的表达调 控有关。在读码框架的5′端,是由起始密码(start codon)AUG开始的,它编码一个蛋氨酸。 在读码框架的3′端,含有一个或一个以上的终止密码(stop codon):UAA、UAG和UGA, 其功能是终止这一多肽链的合成。在真核生物mRNA的3′端,通常还含有转录后加上去的 多聚腺嘌呤核苷酸(polyA)序列作尾巴,其功能可能与增加mRNA分子的稳定性有关。 图12-3 真核生物及原核生物mRNA结构简图 mRNA分子的5′端序列对于起始密码的选择有重要作用,这种作用对于原核生物和真 核生物还有所差别。原核生物中(图12-3),在mRNA分子起始密码子的上游含有一段特殊的 图12-2 正常的与噬菌体T2感染前后大肠杆菌 核糖体的密度梯度超离心分析