正在加载图片...
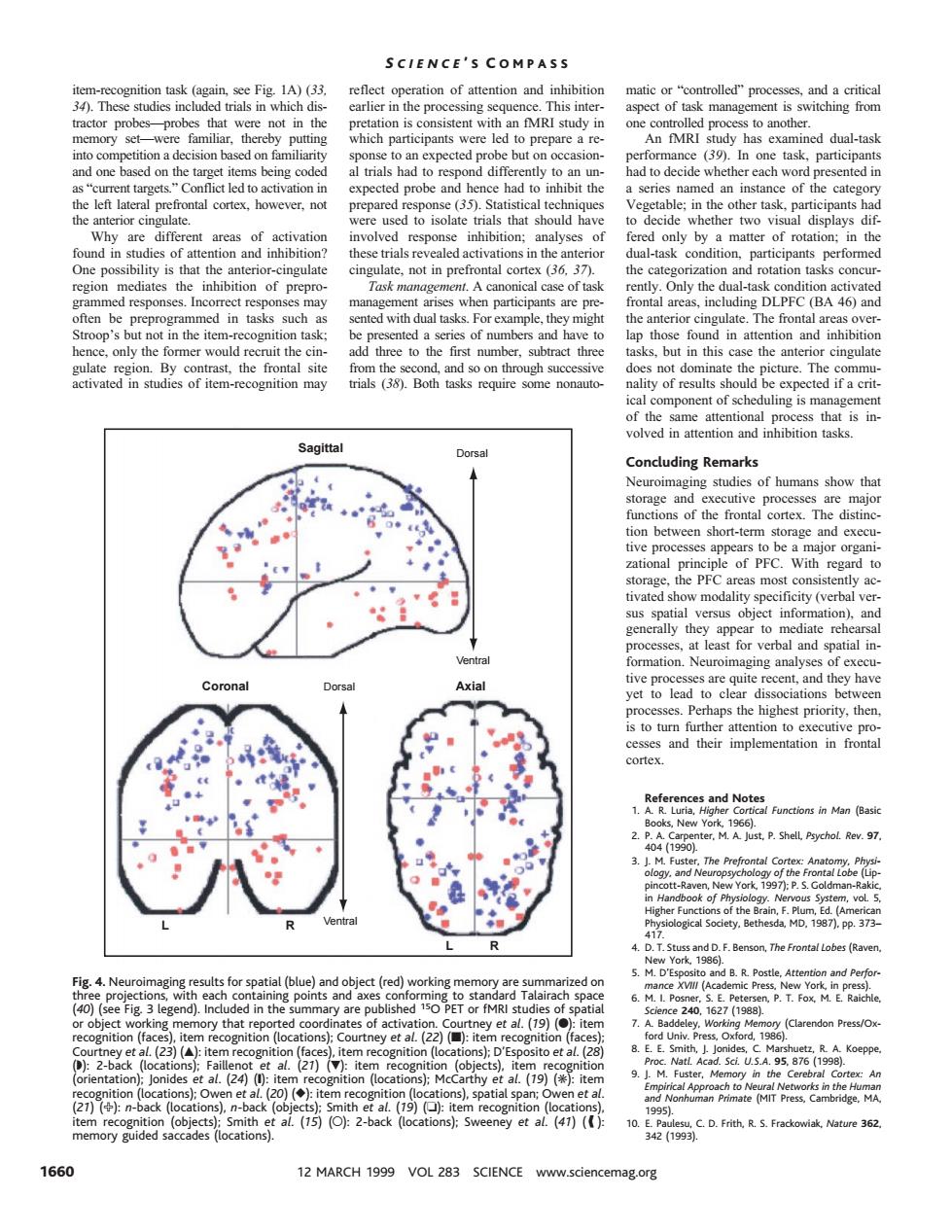
SCIENCE'S COMPASS g sc aspect of task man led to on the (39 of the cater rials that sh oud hav ether two displays di lual-task co ndition participant not in pr 3 he -task conditio ntal Only the (BA 46)an n the iter in atte ntion an and ical nent of scheduline is m Concluding remarks s of the frontal cortex The be PEC the ar to diat Perhaps the highest prority,the cortex otes in Man (Bas 2.P.A Carper .Shell Pychol. 3. The Pre enson.The Frontal Lobes (Rave and B.R.Postle.A ory (Clarendon Press/O 28 2400 s]M (19 ):item nd No temrognaeetotons eney et al(4)( 1660 12 MARCH 1999 VOL 283 SCIENCE www.sciencemag.org item-recognition task (again, see Fig. 1A) (33, 34). These studies included trials in which distractor probes—probes that were not in the memory set—were familiar, thereby putting into competition a decision based on familiarity and one based on the target items being coded as “current targets.” Conflict led to activation in the left lateral prefrontal cortex, however, not the anterior cingulate. Why are different areas of activation found in studies of attention and inhibition? One possibility is that the anterior-cingulate region mediates the inhibition of preprogrammed responses. Incorrect responses may often be preprogrammed in tasks such as Stroop’s but not in the item-recognition task; hence, only the former would recruit the cingulate region. By contrast, the frontal site activated in studies of item-recognition may reflect operation of attention and inhibition earlier in the processing sequence. This interpretation is consistent with an fMRI study in which participants were led to prepare a response to an expected probe but on occasional trials had to respond differently to an unexpected probe and hence had to inhibit the prepared response (35). Statistical techniques were used to isolate trials that should have involved response inhibition; analyses of these trials revealed activations in the anterior cingulate, not in prefrontal cortex (36, 37). Task management. A canonical case of task management arises when participants are presented with dual tasks. For example, they might be presented a series of numbers and have to add three to the first number, subtract three from the second, and so on through successive trials (38). Both tasks require some nonautomatic or “controlled” processes, and a critical aspect of task management is switching from one controlled process to another. An fMRI study has examined dual-task performance (39). In one task, participants had to decide whether each word presented in a series named an instance of the category Vegetable; in the other task, participants had to decide whether two visual displays differed only by a matter of rotation; in the dual-task condition, participants performed the categorization and rotation tasks concurrently. Only the dual-task condition activated frontal areas, including DLPFC (BA 46) and the anterior cingulate. The frontal areas overlap those found in attention and inhibition tasks, but in this case the anterior cingulate does not dominate the picture. The communality of results should be expected if a critical component of scheduling is management of the same attentional process that is involved in attention and inhibition tasks. Concluding Remarks Neuroimaging studies of humans show that storage and executive processes are major functions of the frontal cortex. The distinction between short-term storage and executive processes appears to be a major organizational principle of PFC. With regard to storage, the PFC areas most consistently activated show modality specificity (verbal versus spatial versus object information), and generally they appear to mediate rehearsal processes, at least for verbal and spatial information. Neuroimaging analyses of executive processes are quite recent, and they have yet to lead to clear dissociations between processes. Perhaps the highest priority, then, is to turn further attention to executive processes and their implementation in frontal cortex. References and Notes 1. A. R. Luria, Higher Cortical Functions in Man (Basic Books, New York, 1966). 2. P. A. Carpenter, M. A. Just, P. Shell, Psychol. Rev. 97, 404 (1990). 3. J. M. Fuster, The Prefrontal Cortex: Anatomy, Physiology, and Neuropsychology of the Frontal Lobe (Lippincott-Raven, New York, 1997); P. S. Goldman-Rakic, in Handbook of Physiology. Nervous System, vol. 5, Higher Functions of the Brain, F. Plum, Ed. (American Physiological Society, Bethesda, MD, 1987), pp. 373– 417. 4. D. T. Stuss and D. F. Benson, The Frontal Lobes (Raven, New York, 1986). 5. M. D’Esposito and B. R. Postle, Attention and Performance XVIII (Academic Press, New York, in press). 6. M. I. Posner, S. E. Petersen, P. T. Fox, M. E. Raichle, Science 240, 1627 (1988). 7. A. Baddeley, Working Memory (Clarendon Press/Oxford Univ. Press, Oxford, 1986). 8. E. E. Smith, J. Jonides, C. Marshuetz, R. A. Koeppe, Proc. Natl. Acad. Sci. U.S.A. 95, 876 (1998). 9. J. M. Fuster, Memory in the Cerebral Cortex: An Empirical Approach to Neural Networks in the Human and Nonhuman Primate (MIT Press, Cambridge, MA, 1995). 10. E. Paulesu, C. D. Frith, R. S. Frackowiak, Nature 362, 342 (1993). Sagittal Coronal L L R R Dorsal Axial Ventral Ventral Dorsal Fig. 4. Neuroimaging results for spatial (blue) and object (red) working memory are summarized on three projections, with each containing points and axes conforming to standard Talairach space (40) (see Fig. 3 legend). Included in the summary are published 15O PET or fMRI studies of spatial or object working memory that reported coordinates of activation. Courtney et al. (19) (F): item recognition (faces), item recognition (locations); Courtney et al. (22) (■): item recognition (faces); Courtney et al. (23) (Œ): item recognition (faces), item recognition (locations); D’Esposito et al. (28) (◗): 2-back (locations); Faillenot et al. (21) (): item recognition (objects), item recognition (orientation); Jonides et al. (24) (❙): item recognition (locations); McCarthy et al. (19) (✳): item recognition (locations); Owen et al. (20) (}): item recognition (locations), spatial span; Owen et al. (21) (✙): n-back (locations), n-back (objects); Smith et al. (19) (❏): item recognition (locations), item recognition (objects); Smith et al. (15) (E): 2-back (locations); Sweeney et al. (41) ( ): memory guided saccades (locations). K S CIENCE ’ S C OMPASS 1660 12 MARCH 1999 VOL 283 SCIENCE www.sciencemag.org