正在加载图片...
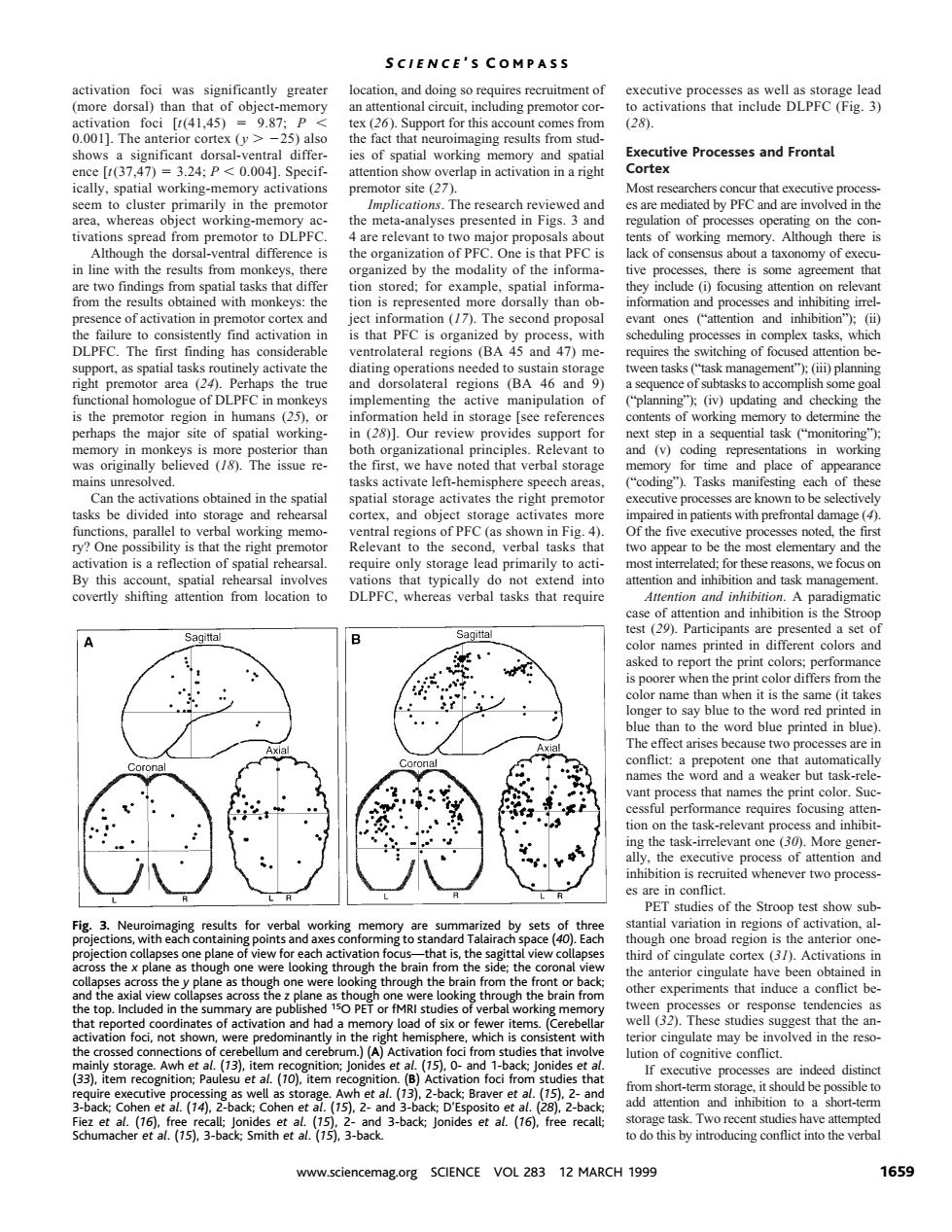
SCIENCE'S COMPASS (28). ho of spatial working Executive Processes and Fronta tial working- ry activations site (27 the met Ca 3a of by PFC and are edby the some ag keys:the dorsally than ob is that PFC is org with ateral reg f to the truc 25 ingd in rage [se refer y to det and (v)c nsks ate left- e speech area coding" Tasks man of the ob ing memo 2 of PFC equire only orage lead primarily to 30m an erba not sh in the res that h et al (D hle CK Braver et at. 3-back lo o do this by int cing conflict into the verb www.sciencemag.org SCIENCE VOL 283 12 MARCH 1999 1659activation foci was significantly greater (more dorsal) than that of object-memory activation foci [t(41,45) 5 9.87; P , 0.001]. The anterior cortex ( y . 225) also shows a significant dorsal-ventral difference [t(37,47) 5 3.24; P , 0.004]. Specifically, spatial working-memory activations seem to cluster primarily in the premotor area, whereas object working-memory activations spread from premotor to DLPFC. Although the dorsal-ventral difference is in line with the results from monkeys, there are two findings from spatial tasks that differ from the results obtained with monkeys: the presence of activation in premotor cortex and the failure to consistently find activation in DLPFC. The first finding has considerable support, as spatial tasks routinely activate the right premotor area (24). Perhaps the true functional homologue of DLPFC in monkeys is the premotor region in humans (25), or perhaps the major site of spatial workingmemory in monkeys is more posterior than was originally believed (18). The issue remains unresolved. Can the activations obtained in the spatial tasks be divided into storage and rehearsal functions, parallel to verbal working memory? One possibility is that the right premotor activation is a reflection of spatial rehearsal. By this account, spatial rehearsal involves covertly shifting attention from location to location, and doing so requires recruitment of an attentional circuit, including premotor cortex (26). Support for this account comes from the fact that neuroimaging results from studies of spatial working memory and spatial attention show overlap in activation in a right premotor site (27). Implications. The research reviewed and the meta-analyses presented in Figs. 3 and 4 are relevant to two major proposals about the organization of PFC. One is that PFC is organized by the modality of the information stored; for example, spatial information is represented more dorsally than object information (17). The second proposal is that PFC is organized by process, with ventrolateral regions (BA 45 and 47) mediating operations needed to sustain storage and dorsolateral regions (BA 46 and 9) implementing the active manipulation of information held in storage [see references in (28)]. Our review provides support for both organizational principles. Relevant to the first, we have noted that verbal storage tasks activate left-hemisphere speech areas, spatial storage activates the right premotor cortex, and object storage activates more ventral regions of PFC (as shown in Fig. 4). Relevant to the second, verbal tasks that require only storage lead primarily to activations that typically do not extend into DLPFC, whereas verbal tasks that require executive processes as well as storage lead to activations that include DLPFC (Fig. 3) (28). Executive Processes and Frontal Cortex Most researchers concur that executive processes are mediated by PFC and are involved in the regulation of processes operating on the contents of working memory. Although there is lack of consensus about a taxonomy of executive processes, there is some agreement that they include (i) focusing attention on relevant information and processes and inhibiting irrelevant ones (“attention and inhibition”); (ii) scheduling processes in complex tasks, which requires the switching of focused attention between tasks (“task management”); (iii) planning a sequence of subtasks to accomplish some goal (“planning”); (iv) updating and checking the contents of working memory to determine the next step in a sequential task (“monitoring”); and (v) coding representations in working memory for time and place of appearance (“coding”). Tasks manifesting each of these executive processes are known to be selectively impaired in patients with prefrontal damage (4). Of the five executive processes noted, the first two appear to be the most elementary and the most interrelated; for these reasons, we focus on attention and inhibition and task management. Attention and inhibition. A paradigmatic case of attention and inhibition is the Stroop test (29). Participants are presented a set of color names printed in different colors and asked to report the print colors; performance is poorer when the print color differs from the color name than when it is the same (it takes longer to say blue to the word red printed in blue than to the word blue printed in blue). The effect arises because two processes are in conflict: a prepotent one that automatically names the word and a weaker but task-relevant process that names the print color. Successful performance requires focusing attention on the task-relevant process and inhibiting the task-irrelevant one (30). More generally, the executive process of attention and inhibition is recruited whenever two processes are in conflict. PET studies of the Stroop test show substantial variation in regions of activation, although one broad region is the anterior onethird of cingulate cortex (31). Activations in the anterior cingulate have been obtained in other experiments that induce a conflict between processes or response tendencies as well (32). These studies suggest that the anterior cingulate may be involved in the resolution of cognitive conflict. If executive processes are indeed distinct from short-term storage, it should be possible to add attention and inhibition to a short-term storage task. Two recent studies have attempted to do this by introducing conflict into the verbal Fig. 3. Neuroimaging results for verbal working memory are summarized by sets of three projections, with each containing points and axes conforming to standard Talairach space (40). Each projection collapses one plane of view for each activation focus—that is, the sagittal view collapses across the x plane as though one were looking through the brain from the side; the coronal view collapses across the y plane as though one were looking through the brain from the front or back; and the axial view collapses across the z plane as though one were looking through the brain from the top. Included in the summary are published 15O PET or fMRI studies of verbal working memory that reported coordinates of activation and had a memory load of six or fewer items. (Cerebellar activation foci, not shown, were predominantly in the right hemisphere, which is consistent with the crossed connections of cerebellum and cerebrum.) (A) Activation foci from studies that involve mainly storage. Awh et al. (13), item recognition; Jonides et al. (15), 0- and 1-back; Jonides et al. (33), item recognition; Paulesu et al. (10), item recognition. (B) Activation foci from studies that require executive processing as well as storage. Awh et al. (13), 2-back; Braver et al. (15), 2- and 3-back; Cohen et al. (14), 2-back; Cohen et al. (15), 2- and 3-back; D’Esposito et al. (28), 2-back; Fiez et al. (16), free recall; Jonides et al. (15), 2- and 3-back; Jonides et al. (16), free recall; Schumacher et al. (15), 3-back; Smith et al. (15), 3-back. S CIENCE ’ S C OMPASS www.sciencemag.org SCIENCE VOL 283 12 MARCH 1999 1659