正在加载图片...
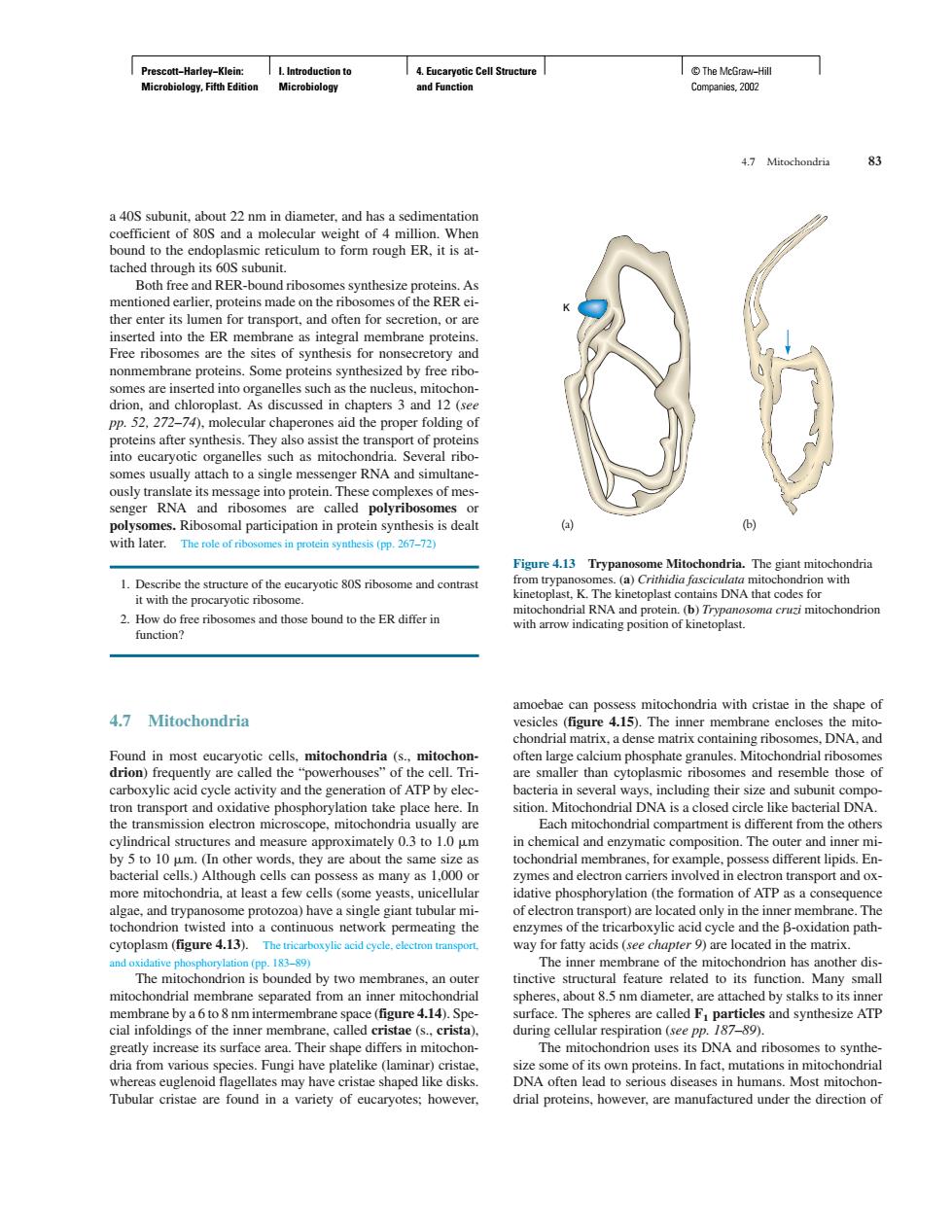
Cal Both free and RER-bound ribos mbrane protein or non ory an 270cc tof pro omes usually attach to a single messenger RNA and simultane mes.Ribo omal participation n protein synthesis is dealt (b) with later.Thef mes in protein synthesis (pp.267-72) omes and those bound to the ER differ in s mitochondria with cristae in the shane of 4.7 Mitochondria Found in most cre called ium phosphate ondrial rib es and res e o eon m sition.Mitoch I DNA ope,mito the others ranes,for example,poss s different lipic En of electron ytoplasm (figure 4.13).The mitochondrial membrane separated from an iner mito eres.about 5 pm diameter are attac ed by stalks to its inne are 4.14).Spe ace. es and synthesize ATP Tubular cristae are found in a variety of eucaryotes:however. drial proteins,however,are manufactured under the direction of Prescott−Harley−Klein: Microbiology, Fifth Edition I. Introduction to Microbiology 4. Eucaryotic Cell Structure and Function © The McGraw−Hill Companies, 2002 a 40S subunit, about 22 nm in diameter, and has a sedimentation coefficient of 80S and a molecular weight of 4 million. When bound to the endoplasmic reticulum to form rough ER, it is attached through its 60S subunit. Both free and RER-bound ribosomes synthesize proteins. As mentioned earlier, proteins made on the ribosomes of the RER either enter its lumen for transport, and often for secretion, or are inserted into the ER membrane as integral membrane proteins. Free ribosomes are the sites of synthesis for nonsecretory and nonmembrane proteins. Some proteins synthesized by free ribosomes are inserted into organelles such as the nucleus, mitochondrion, and chloroplast. As discussed in chapters 3 and 12 (see pp. 52, 272–74), molecular chaperones aid the proper folding of proteins after synthesis. They also assist the transport of proteins into eucaryotic organelles such as mitochondria. Several ribosomes usually attach to a single messenger RNA and simultaneously translate its message into protein. These complexes of messenger RNA and ribosomes are called polyribosomes or polysomes. Ribosomal participation in protein synthesis is dealt with later. The role of ribosomes in protein synthesis (pp. 267–72) 1. Describe the structure of the eucaryotic 80S ribosome and contrast it with the procaryotic ribosome. 2. How do free ribosomes and those bound to the ER differ in function? 4.7 Mitochondria Found in most eucaryotic cells, mitochondria (s., mitochondrion) frequently are called the “powerhouses” of the cell. Tricarboxylic acid cycle activity and the generation of ATP by electron transport and oxidative phosphorylation take place here. In the transmission electron microscope, mitochondria usually are cylindrical structures and measure approximately 0.3 to 1.0 m by 5 to 10 m. (In other words, they are about the same size as bacterial cells.) Although cells can possess as many as 1,000 or more mitochondria, at least a few cells (some yeasts, unicellular algae, and trypanosome protozoa) have a single giant tubular mitochondrion twisted into a continuous network permeating the cytoplasm (figure 4.13). The tricarboxylic acid cycle, electron transport, and oxidative phosphorylation (pp. 183–89) The mitochondrion is bounded by two membranes, an outer mitochondrial membrane separated from an inner mitochondrial membrane by a 6 to 8 nm intermembrane space (figure 4.14). Special infoldings of the inner membrane, called cristae (s., crista), greatly increase its surface area. Their shape differs in mitochondria from various species. Fungi have platelike (laminar) cristae, whereas euglenoid flagellates may have cristae shaped like disks. Tubular cristae are found in a variety of eucaryotes; however, amoebae can possess mitochondria with cristae in the shape of vesicles (figure 4.15). The inner membrane encloses the mitochondrial matrix, a dense matrix containing ribosomes, DNA, and often large calcium phosphate granules. Mitochondrial ribosomes are smaller than cytoplasmic ribosomes and resemble those of bacteria in several ways, including their size and subunit composition. Mitochondrial DNA is a closed circle like bacterial DNA. Each mitochondrial compartment is different from the others in chemical and enzymatic composition. The outer and inner mitochondrial membranes, for example, possess different lipids. Enzymes and electron carriers involved in electron transport and oxidative phosphorylation (the formation of ATP as a consequence of electron transport) are located only in the inner membrane. The enzymes of the tricarboxylic acid cycle and the -oxidation pathway for fatty acids (see chapter 9) are located in the matrix. The inner membrane of the mitochondrion has another distinctive structural feature related to its function. Many small spheres, about 8.5 nm diameter, are attached by stalks to its inner surface. The spheres are called F1 particles and synthesize ATP during cellular respiration (see pp. 187–89). The mitochondrion uses its DNA and ribosomes to synthesize some of its own proteins. In fact, mutations in mitochondrial DNA often lead to serious diseases in humans. Most mitochondrial proteins, however, are manufactured under the direction of 4.7 Mitochondria 83 K Figure 4.13 Trypanosome Mitochondria. The giant mitochondria from trypanosomes. (a) Crithidia fasciculata mitochondrion with kinetoplast, K. The kinetoplast contains DNA that codes for mitochondrial RNA and protein. (b) Trypanosoma cruzi mitochondrion with arrow indicating position of kinetoplast. (a) (b)��