正在加载图片...
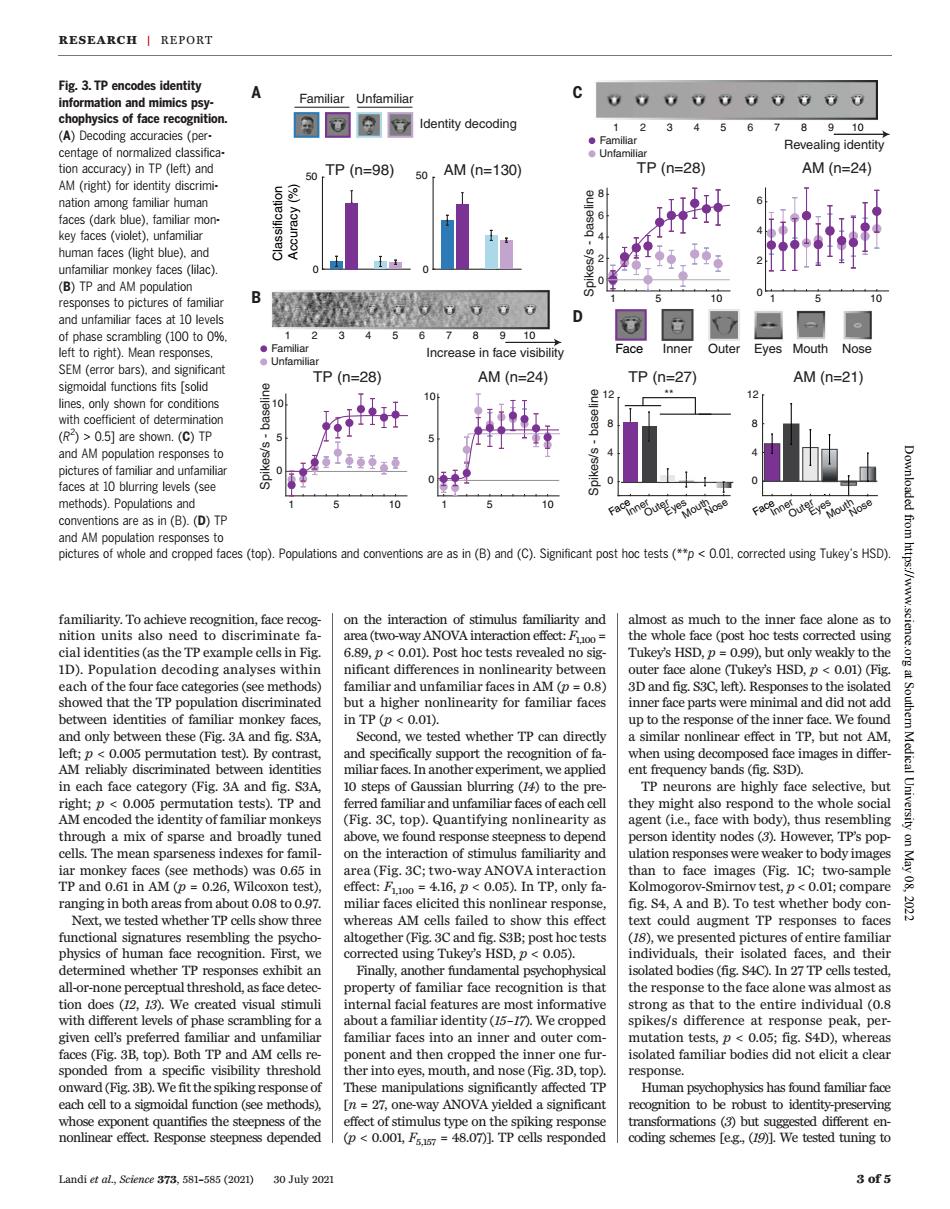
RESEARCH I REPORT Fig 3 IP encodes identity A information and mimics psy 0000000000 (A)Decoding accuracies (pe a同a7 ldentity decoding 12345678910 Revealing identity n )()nd 0,TP(n=98)50,AM(n=130 TP(n=28) AM(n=24】 是8 (B)TP and AM populatio 0 eft t)( 34 回回可 Increase in face visibility Face Inner Outer Eyes Mouth os sav(( an TP(n=28) AM(n=24) TP(n=27) AM(n=21) (C)TP es to 10 (D)TP picturesofwhole and cropped faces(top).Populations and conventions are as in (B)and (C).Significant post ho tests(<using Tukey's HSD) nition,face reco on the interactior of stimulus familiarit and much to the inner face alone a on ef tests correc 001) lad D).Population decoding analyses ithi nificant differe er face ald e (Tukey's HSD,p<0.01)(Fi in TP (p<0.01). up to the response of the inner face.We found rt the ition of ing de d fa AM D d een n anoth en requency ban familiar and unfamiliar fac of each thev might also resp nd to the whole ugh a mix of )Quantifying nonline ty a y),thus re identity nodes TP's on the inter ion of stimulus familiarity tio responses were weaker to body image IP and AM 02. 416. 0 May 08, ranging in both eicitet ng.S4, Aand B).To twhether bodyo nal signa atures resemb the her (Fig 3C S3B:D est ()we prese s of entire familiar the m l-o roperty of familiar face recognition is tha as almost as e peak vencel's preferred fam ar a nilia s into an ir er and outer m t a clea hold mouth,and nose (D.top Fig.3B nipulat stonmatio (3)but 30Juy202 familiarity. To achieve recognition, face recognition units also need to discriminate facial identities (as the TP example cells in Fig. 1D). Population decoding analyses within each of the four face categories (see methods) showed that the TP population discriminated between identities of familiar monkey faces, and only between these (Fig. 3A and fig. S3A, left; p < 0.005 permutation test). By contrast, AM reliably discriminated between identities in each face category (Fig. 3A and fig. S3A, right; p < 0.005 permutation tests). TP and AM encoded the identity of familiar monkeys through a mix of sparse and broadly tuned cells. The mean sparseness indexes for familiar monkey faces (see methods) was 0.65 in TP and 0.61 in AM (p = 0.26, Wilcoxon test), ranging in both areas from about 0.08 to 0.97. Next, we tested whether TP cells show three functional signatures resembling the psychophysics of human face recognition. First, we determined whether TP responses exhibit an all-or-none perceptual threshold, as face detection does (12, 13). We created visual stimuli with different levels of phase scrambling for a given cell’s preferred familiar and unfamiliar faces (Fig. 3B, top). Both TP and AM cells responded from a specific visibility threshold onward (Fig. 3B). We fit the spiking response of each cell to a sigmoidal function (see methods), whose exponent quantifies the steepness of the nonlinear effect. Response steepness depended on the interaction of stimulus familiarity and area (two-way ANOVA interaction effect: F1,100 = 6.89, p < 0.01). Post hoc tests revealed no significant differences in nonlinearity between familiar and unfamiliar faces in AM (p = 0.8) but a higher nonlinearity for familiar faces in TP (p < 0.01). Second, we tested whether TP can directly and specifically support the recognition of familiar faces. In another experiment, we applied 10 steps of Gaussian blurring (14) to the preferred familiar and unfamiliar faces of each cell (Fig. 3C, top). Quantifying nonlinearity as above, we found response steepness to depend on the interaction of stimulus familiarity and area (Fig. 3C; two-way ANOVA interaction effect: F1,100 = 4.16, p < 0.05). In TP, only familiar faces elicited this nonlinear response, whereas AM cells failed to show this effect altogether (Fig. 3C and fig. S3B; post hoc tests corrected using Tukey’s HSD, p < 0.05). Finally, another fundamental psychophysical property of familiar face recognition is that internal facial features are most informative about a familiar identity (15–17). We cropped familiar faces into an inner and outer component and then cropped the inner one further into eyes, mouth, and nose (Fig. 3D, top). These manipulations significantly affected TP [n = 27, one-way ANOVA yielded a significant effect of stimulus type on the spiking response (p < 0.001, F5,157 = 48.07)]. TP cells responded almost as much to the inner face alone as to the whole face (post hoc tests corrected using Tukey’s HSD, p = 0.99), but only weakly to the outer face alone (Tukey’s HSD, p < 0.01) (Fig. 3D and fig. S3C, left). Responses to the isolated inner face parts were minimal and did not add up to the response of the inner face. We found a similar nonlinear effect in TP, but not AM, when using decomposed face images in different frequency bands (fig. S3D). TP neurons are highly face selective, but they might also respond to the whole social agent (i.e., face with body), thus resembling person identity nodes (3). However, TP’s population responses were weaker to body images than to face images (Fig. 1C; two-sample Kolmogorov-Smirnov test, p < 0.01; compare fig. S4, A and B). To test whether body context could augment TP responses to faces (18), we presented pictures of entire familiar individuals, their isolated faces, and their isolated bodies (fig. S4C). In 27 TP cells tested, the response to the face alone was almost as strong as that to the entire individual (0.8 spikes/s difference at response peak, permutation tests, p < 0.05; fig. S4D), whereas isolated familiar bodies did not elicit a clear response. Human psychophysics has found familiar face recognition to be robust to identity-preserving transformations (3) but suggested different encoding schemes [e.g., (19)]. We tested tuning to Landi et al., Science 373, 581–585 (2021) 30 July 2021 3 of 5 Fig. 3. TP encodes identity information and mimics psychophysics of face recognition. (A) Decoding accuracies (percentage of normalized classification accuracy) in TP (left) and AM (right) for identity discrimination among familiar human faces (dark blue), familiar monkey faces (violet), unfamiliar human faces (light blue), and unfamiliar monkey faces (lilac). (B) TP and AM population responses to pictures of familiar and unfamiliar faces at 10 levels of phase scrambling (100 to 0%, left to right). Mean responses, SEM (error bars), and significant sigmoidal functions fits [solid lines, only shown for conditions with coefficient of determination (R2 ) > 0.5] are shown. (C) TP and AM population responses to pictures of familiar and unfamiliar faces at 10 blurring levels (see methods). Populations and conventions are as in (B). (D) TP and AM population responses to pictures of whole and cropped faces (top). Populations and conventions are as in (B) and (C). Significant post hoc tests (**p < 0.01, corrected using Tukey’s HSD). A B TP (n=28) AM (n=24) 0 5 10 Spikes/s - baseline 0 5 10 0 50 0 50 Classification Accuracy (%) 0 4 8 12 Spikes/s - baseline ** TP (n=27) AM (n=21) 0 4 8 12 TP (n=98) AM (n=130) Identity decoding C 0 2 4 6 Spikes/s - baseline 0 2 4 6 8 TP (n=28) AM (n=24) Face Inner Eyes Mouth Nose Outer D Familiar Unfamiliar Increase in face visibility Familiar Unfamiliar 1 2 345 6 7 8 9 10 1 5 10 1 5 10 Revealing identity Familiar Unfamiliar 1 2 345 6 7 8 9 10 1 5 10 1 5 10 FaceInner OuterEyes MouthNose FaceInner OuterEyes MouthNose RESEARCH | REPORT Downloaded from https://www.science.org at Southern Medical University on May 08, 2022