正在加载图片...
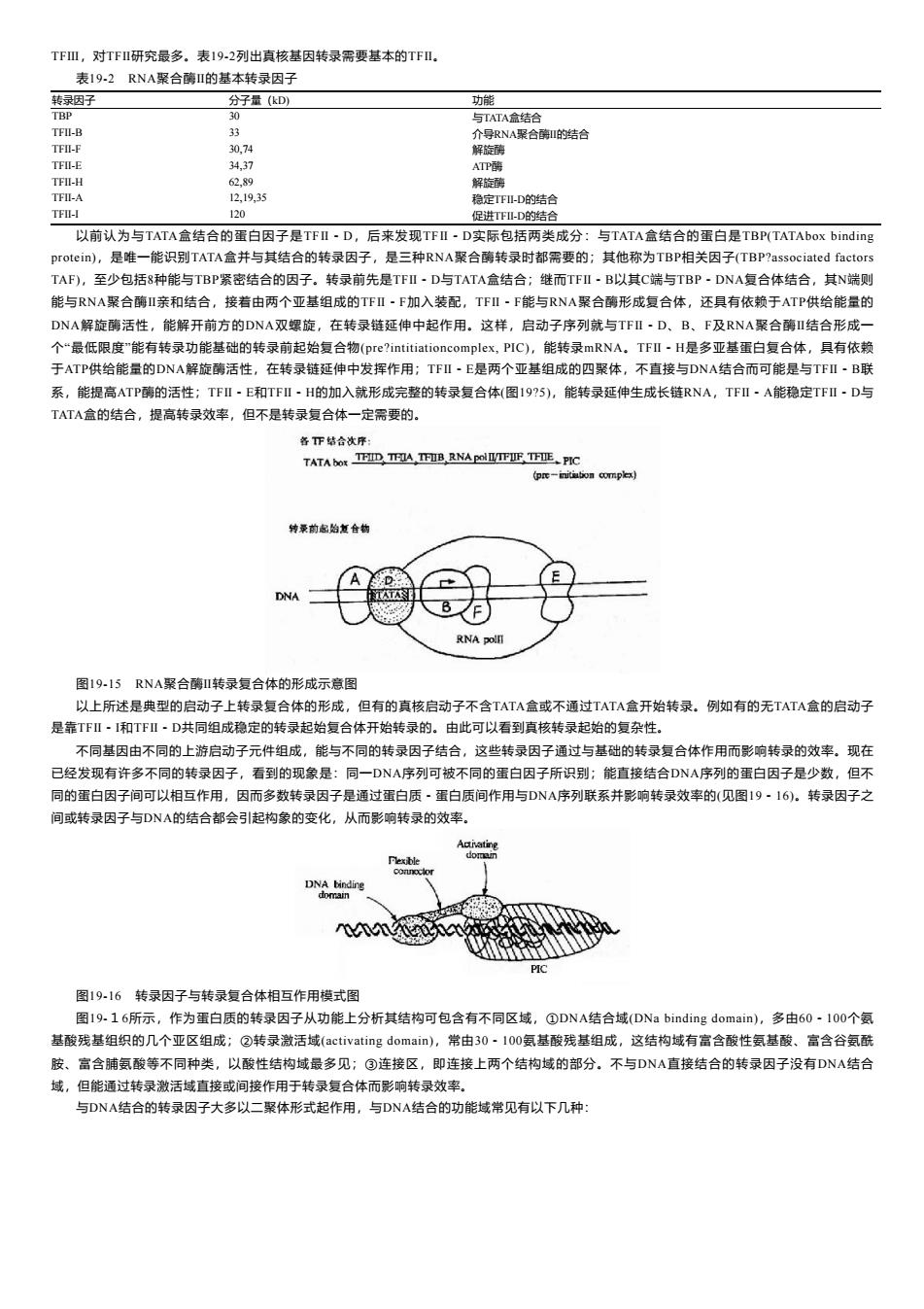
TFⅢ,对TFⅡ研究最多。表192列出真核基因转录需要基本的TFⅡ. 表19-2RNA聚合酶I的基本转录因子 转录因子 分子量(kD) 功能 TBP 30 与TATA盒结合 TFI-B 33 介导RNA聚合酶的结合 TFI-F 30,74 解旋酶 TFI-E 34,37 ATP薛 TFII-H 62,89 解旋藓 TFII-A 12,19,35 稳定TFI-D的结合 TFI-I 120 促进TF-D的结合 以前认为与TATA盒结合的蛋白因子是TFⅡ~D,后来发现TFⅡ·D实际包括两类成分:与TATA盒结合的蛋白是TBP(TATAbox binding protein),是唯一能识别TATA盒并与其结合的转录因子,是三种RNA聚合酶转录时都需要的;其他称为TBP相关因子(TBP?associated factors TAF),至少包括8种能与TBP紧密结合的因子。转录前先是TFI·D与TATA盒结合;继而TFI·B以其C端与TBP,DNA复合体结合,其N端则 能与RNA聚合酶I亲和结合,接着由两个亚基组成的TFI·F加入装配,TFI·F能与RNA聚合酶形成复合体,还具有依赖于ATP供给能量的 DNA解旋酶活性,能解开前方的DNA双螺旋,在转录链延伸中起作用。这样,启动子序列就与TFⅡ·D、B、F及RNA聚合酶Ⅱ结合形成一 个“最低限度"能有转录功能基础的转录前起始复合物(pre?intitiationcomplex,PIC),能转录mRNA。TFI~H是多亚基蛋白复合体,具有依赖 于ATP供给能量的DNA解旋酶活性,在转录链延伸中发挥作用;TFI·E是两个亚基组成的四聚体,不直接与DNA结合而可能是与TFⅡ-B联 系,能提高ATP酶的活性;TFⅡ-E和TFⅡ-H的加入就形成完整的转录复合体(图19?5),能转录延伸生成长链RNA,TFⅡ·A能稳定TFⅡ·D与 TATA盒的结合,提高转录效率,但不是转录复合体一定需要的。 各T下结合次序: TATA bOR TF四,TFA,TB,RNAPOTFUF TFI正、PIC (pre-initiution complex) 特桑前虚始复合物 DNA RNA pollT 图19-15RNA聚合酶I转录复合体的形成示意图 以上所述是典型的启动子上转录复合体的形成,但有的真核启动子不含TATA盒或不通过TATA盒开始转录。例如有的无TATA盒的启动子 是靠T℉Ⅱ·I和T℉Ⅱ·D共同组成稳定的转录起始复合体开始转录的。由此可以看到真核转录起始的复杂性。 不同基因由不同的上游启动子元件组成,能与不同的转录因子结合,这些转录因子通过与基础的转录复合体作用而影响转录的效率。现在 已经发现有许多不同的转录因子,看到的现象是:同一DNA序列可被不同的蛋白因子所识别:能直接结合DNA序列的蛋白因子是少数,但不 同的蛋白因子间可以相互作用,因而多数转录因子是通过蛋白质·蛋白质间作用与DNA序列联系并影响转录效率的(见图19·16)。转录因子之 间或转录因子与DNA的结合都会引起构象的变化,从而影响转录的效率。 Flexible DNA binding domain 图19-16转录因子与转录复合体相互作用模式图 图19.16所示,作为蛋白质的转录因子从功能上分析其结构可包含有不同区域,①DNA结合域(DNa binding domain),多由60·100个氨 基酸残基组织的几个亚区组成;②转录激活域(activating domain),常由30·I00氨基酸残基组成,这结构域有富含酸性氨基酸、富含谷氨酰 胺、富含脯氨酸等不同种类,以酸性结构域最多见;③连接区,即连接上两个结构域的部分。不与DNA直接结合的转录因子没有DNA结合 域,但能通过转录激活域直接或间接作用于转录复合体而影响转录效率。 与DNA结合的转录因子大多以二聚体形式起作用,与DNA结合的功能域常见有以下几种:TFⅢ,对TFⅡ研究最多。表19-2列出真核基因转录需要基本的TFⅡ。 表19-2 RNA聚合酶Ⅱ的基本转录因子 转录因子 分子量(kD) 功能 TBP 30 与TATA盒结合 TFⅡ-B 33 介导RNA聚合酶Ⅱ的结合 TFⅡ-F 30,74 解旋酶 TFⅡ-E 34,37 ATP酶 TFⅡ-H 62,89 解旋酶 TFⅡ-A 12,19,35 稳定TFⅡ-D的结合 TFⅡ-I 120 促进TFⅡ-D的结合 以前认为与TATA盒结合的蛋白因子是TFⅡ-D,后来发现TFⅡ-D实际包括两类成分:与TATA盒结合的蛋白是TBP(TATAbox binding protein),是唯一能识别TATA盒并与其结合的转录因子,是三种RNA聚合酶转录时都需要的;其他称为TBP相关因子(TBP?associated factors TAF),至少包括8种能与TBP紧密结合的因子。转录前先是TFⅡ-D与TATA盒结合;继而TFⅡ-B以其C端与TBP-DNA复合体结合,其N端则 能与RNA聚合酶Ⅱ亲和结合,接着由两个亚基组成的TFⅡ-F加入装配,TFⅡ-F能与RNA聚合酶形成复合体,还具有依赖于ATP供给能量的 DNA解旋酶活性,能解开前方的DNA双螺旋,在转录链延伸中起作用。这样,启动子序列就与TFⅡ-D、B、F及RNA聚合酶Ⅱ结合形成一 个“最低限度”能有转录功能基础的转录前起始复合物(pre?intitiationcomplex, PIC),能转录mRNA。TFⅡ-H是多亚基蛋白复合体,具有依赖 于ATP供给能量的DNA解旋酶活性,在转录链延伸中发挥作用;TFⅡ-E是两个亚基组成的四聚体,不直接与DNA结合而可能是与TFⅡ-B联 系,能提高ATP酶的活性;TFⅡ-E和TFⅡ-H的加入就形成完整的转录复合体(图19?5),能转录延伸生成长链RNA,TFⅡ-A能稳定TFⅡ-D与 TATA盒的结合,提高转录效率,但不是转录复合体一定需要的。 图19-15 RNA聚合酶Ⅱ转录复合体的形成示意图 以上所述是典型的启动子上转录复合体的形成,但有的真核启动子不含TATA盒或不通过TATA盒开始转录。例如有的无TATA盒的启动子 是靠TFⅡ-I和TFⅡ-D共同组成稳定的转录起始复合体开始转录的。由此可以看到真核转录起始的复杂性。 不同基因由不同的上游启动子元件组成,能与不同的转录因子结合,这些转录因子通过与基础的转录复合体作用而影响转录的效率。现在 已经发现有许多不同的转录因子,看到的现象是:同一DNA序列可被不同的蛋白因子所识别;能直接结合DNA序列的蛋白因子是少数,但不 同的蛋白因子间可以相互作用,因而多数转录因子是通过蛋白质-蛋白质间作用与DNA序列联系并影响转录效率的(见图19-16)。转录因子之 间或转录因子与DNA的结合都会引起构象的变化,从而影响转录的效率。 图19-16 转录因子与转录复合体相互作用模式图 图19-16所示,作为蛋白质的转录因子从功能上分析其结构可包含有不同区域,①DNA结合域(DNa binding domain),多由60-100个氨 基酸残基组织的几个亚区组成;②转录激活域(activating domain),常由30-100氨基酸残基组成,这结构域有富含酸性氨基酸、富含谷氨酰 胺、富含脯氨酸等不同种类,以酸性结构域最多见;③连接区,即连接上两个结构域的部分。不与DNA直接结合的转录因子没有DNA结合 域,但能通过转录激活域直接或间接作用于转录复合体而影响转录效率。 与DNA结合的转录因子大多以二聚体形式起作用,与DNA结合的功能域常见有以下几种: