正在加载图片...
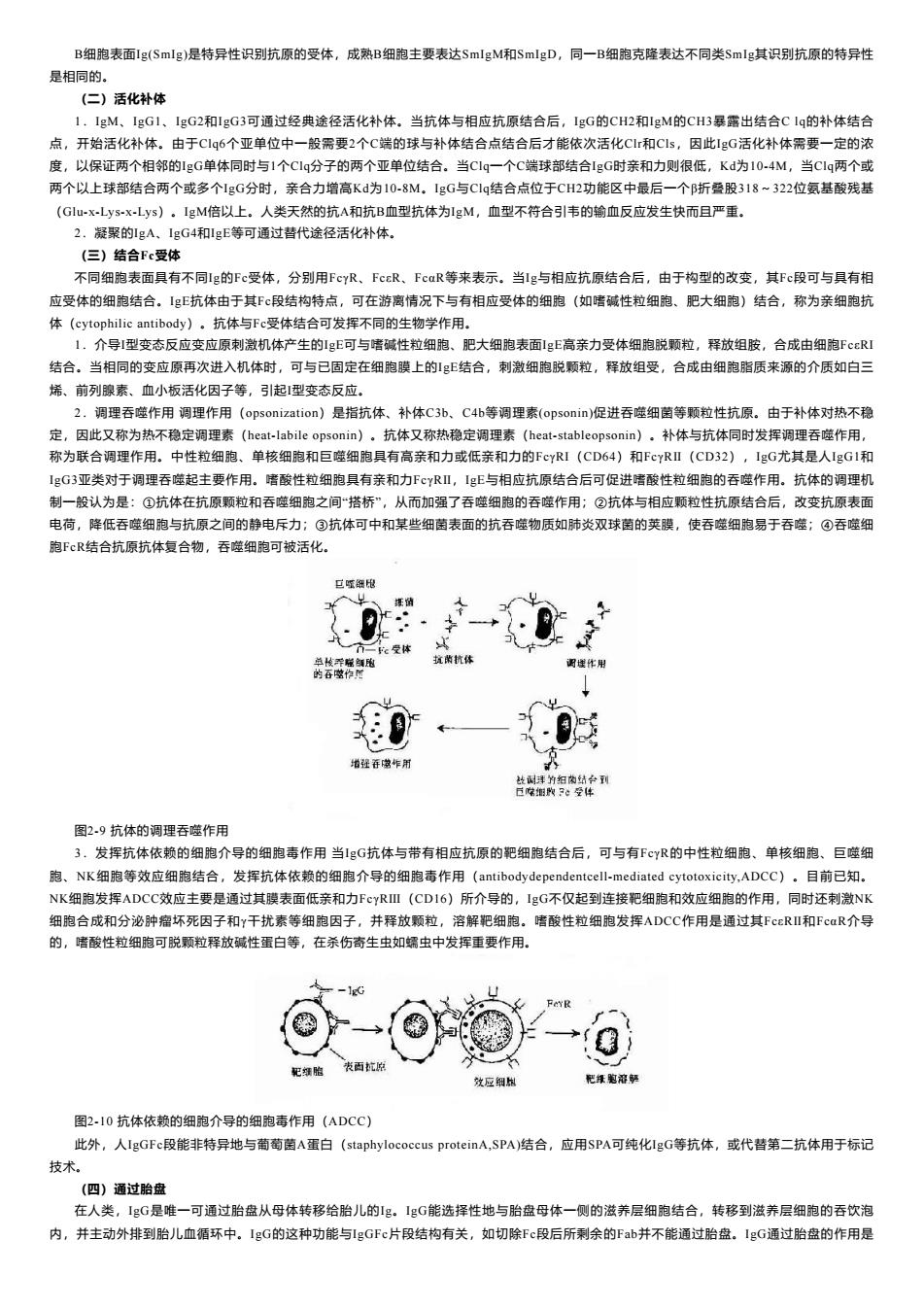
B细胞表面1g(Smlg)是特异性识别抗原的受体,成熟B细跑主要表达SmlgM和SmIgD,同一B细胞克降表达不同类Smlg其识别抗原的特异性 是相同的。 (二)活化补体 1,lgM,lgG1,1gG2和1gG3可通过经典途径活化补体.当抗体与相应抗原结合后,lgG的CH2和gM的CH3暴露出结合Clq的补体结合 点,开始活化补体,由于C1q6个亚单位中一般需要2个C端的球与补体结合点结合后才能依次活化C和Cs,因此1gG活化补体需要一定的浓 度,以保证两个相邻的gG单体同时与1个C1g分子的两个亚单位结合。当CIg一个C端球部结合1gG时亲和力则很低,Kd为10-4M,当C1q两个或 两个以上球部结合两个或多个IgG分时,亲合力增高Kd为10-8M.1eG与C1g结合点位于CH2功能区中最后一个折叠股318~322位氨基酸残基 (G-xLys-xLys)。lgM倍以上,人类天然的抗A和抗B血型抗体为1gM,血型不符合引韦的输血反应发生快而且严重. 2.凝聚的gA、1gG4和gE等可通过皆代途径活化补体. (但)结合Fc受体 不同细跑表而且有不同Ig的Fc受体,分别用FGYR,FccR、。FcR等来表示,当Ig与相应抗原结合后,由于构型的改变,其Fc段可与且有相 应受体的细胞结合。.g抗体由于其F©段结构特点,可在游离情况下与有相应受体的细孢(如储碱性粒细胞、肥大细胞)结合,称为亲细胞抗 体(cytophilic antibody),抗体与Fc受体结合可发挥不同的生物学作用. 1.介导引型变态反应变应原刺激机体产生的1gE可与嗜碱性粒细胞.肥大细胞表面gE高亲力受体细胞脱颗粒,释放组胺,合成由细胞F©RI 结合。当相同的变应原再次进入机体时,可与已固定在细跑膜上的多结合,刺激细陶脱颗粒,释放组受,合成由细胞脂质来源的介质如白三 烯、前列腺素、血小板活化因子等,引起型变态反应。 2.调理吞噬作用调理作用(op5 onization)是指抗体、补体C3b.C4b等调理素(opson1:m)促进吞噬细菌等颗粒性抗原.由于补体对热不稳 定,因此又称为热不稳定调理素(hcat-labile opsonin)。抗体又称热稳定调理素(hcat-stableopsonin),补体与抗体同时发挥调理吞德作用。 称为联合调理作用。中性粒细孢、单核细胞和巨莲细胞具有高亲和力或低亲和力的FcYRI(CD64)和FCYRⅡ(CD32),lgG尤其是人IgG1和 1eG3亚类对于调理吞噬起主要作用。墙酸性粒细胞具有亲和力FyRⅡ,1gE与相应抗原结合后可促进嗜酸性粒细跑的吞噬作用。抗体的调理机 制一般认为是:©抗体在抗原颗粒和吞壁细胞之间搭桥”,从而如强了吞壁细胞的吞壁作用:②抗体与相应颗位性抗原结合后,改变抗原表面 电荷,降低吞噬细跑与抗原之间的静电斥力:③抗体可中和某些细菌表面的抗吞噬物质如肺炎双球萄的英膜,使吞噬细胞易于吞噬:©吞噬细 胞FcR结合抗原抗体复合物,吞噬细跑可被活化, 0· {0 兰为包快经性 图29抗体的调理吞噬作用 发挥抗体依赖的细胞介导的细胞毒作用当G抗体与带有相应抗原的配细胞结合后,可与有F©yR的中性粒细胞、单核细胞。巨噬年 胞.NK细孢等效应细胞结合,发挥抗体依赖的细胞介导的细孢毒作用(antibodydependent-mediated ytotoxiciy.ADCC),目前已知, NK细胞发挥ADCC效应主要是通过其膜表面低亲和力FcyRⅢ(CDI6)所介导的,IgG不仅起到连接纪细孢和效应细胞的作用,同时还刺激NK 细胞合成和分论肿岩坏死因子和Y干扰素等细泡因子,并释放颗粒,溶解把细跑。嗜酸性粒细胞发挥ADCC作用是通过其FcRⅢ和FcaR介导 的,墙酸性粒细胞可脱颗粒释放碱性蛋白等,在杀伤寄生虫如蠕虫中发挥重要作用 图2.10抗体依赖的细跑介导的细胞声作用(ADCC) 此外,人IgGFe段能非特异地与萄萄菌A蛋白((staphylococcus proteinA.SPA)结合,应用SPA可纯化IgG等抗体,或代替第二抗体用于标记 技术 (四)通过胎盘 在人类,1gG是雅一可通过胎盘从母体转移给胎儿的g,1gG能选择性地与胎盘母体一侧的滋养层细胞结合,转移到滋养层细胞的吞饮泡 内,并主动外排到胎儿血须环中,1gG的这种功能与gGFc片段结构有关,如切除Fc段后所利余的Fab并不能通过胎盘。1gG通过胎盘的作用是B细胞表面Ig(SmIg)是特异性识别抗原的受体,成熟B细胞主要表达SmIgM和SmIgD,同一B细胞克隆表达不同类SmIg其识别抗原的特异性 是相同的。 (二)活化补体 1.IgM、IgG1、IgG2和IgG3可通过经典途径活化补体。当抗体与相应抗原结合后,IgG的CH2和IgM的CH3暴露出结合C lq的补体结合 点,开始活化补体。由于Clq6个亚单位中一般需要2个C端的球与补体结合点结合后才能依次活化Clr和Cls,因此IgG活化补体需要一定的浓 度,以保证两个相邻的IgG单体同时与1个Clq分子的两个亚单位结合。当Clq一个C端球部结合IgG时亲和力则很低,Kd为10-4M,当Clq两个或 两个以上球部结合两个或多个IgG分时,亲合力增高Kd为10-8M。IgG与Clq结合点位于CH2功能区中最后一个β折叠股318~322位氨基酸残基 (Glu-x-Lys-x-Lys)。IgM倍以上。人类天然的抗A和抗B血型抗体为IgM,血型不符合引韦的输血反应发生快而且严重。 2.凝聚的IgA、IgG4和IgE等可通过替代途径活化补体。 (三)结合Fc受体 不同细胞表面具有不同Ig的Fc受体,分别用FcγR、FcεR、FcαR等来表示。当Ig与相应抗原结合后,由于构型的改变,其Fc段可与具有相 应受体的细胞结合。IgE抗体由于其Fc段结构特点,可在游离情况下与有相应受体的细胞(如嗜碱性粒细胞、肥大细胞)结合,称为亲细胞抗 体(cytophilic antibody)。抗体与Fc受体结合可发挥不同的生物学作用。 1.介导I型变态反应变应原刺激机体产生的IgE可与嗜碱性粒细胞、肥大细胞表面IgE高亲力受体细胞脱颗粒,释放组胺,合成由细胞FcεRI 结合。当相同的变应原再次进入机体时,可与已固定在细胞膜上的IgE结合,刺激细胞脱颗粒,释放组受,合成由细胞脂质来源的介质如白三 烯、前列腺素、血小板活化因子等,引起Ⅰ型变态反应。 2.调理吞噬作用 调理作用(opsonization)是指抗体、补体C3b、C4b等调理素(opsonin)促进吞噬细菌等颗粒性抗原。由于补体对热不稳 定,因此又称为热不稳定调理素(heat-labile opsonin)。抗体又称热稳定调理素(heat-stableopsonin)。补体与抗体同时发挥调理吞噬作用, 称为联合调理作用。中性粒细胞、单核细胞和巨噬细胞具有高亲和力或低亲和力的FcγRI(CD64)和FcγRⅡ(CD32),IgG尤其是人IgG1和 IgG3亚类对于调理吞噬起主要作用。嗜酸性粒细胞具有亲和力FcγRⅡ,IgE与相应抗原结合后可促进嗜酸性粒细胞的吞噬作用。抗体的调理机 制一般认为是:①抗体在抗原颗粒和吞噬细胞之间“搭桥”,从而加强了吞噬细胞的吞噬作用;②抗体与相应颗粒性抗原结合后,改变抗原表面 电荷,降低吞噬细胞与抗原之间的静电斥力;③抗体可中和某些细菌表面的抗吞噬物质如肺炎双球菌的荚膜,使吞噬细胞易于吞噬;④吞噬细 胞FcR结合抗原抗体复合物,吞噬细胞可被活化。 图2-9 抗体的调理吞噬作用 3.发挥抗体依赖的细胞介导的细胞毒作用 当IgG抗体与带有相应抗原的靶细胞结合后,可与有FcγR的中性粒细胞、单核细胞、巨噬细 胞、NK细胞等效应细胞结合,发挥抗体依赖的细胞介导的细胞毒作用(antibodydependentcell-mediated cytotoxicity,ADCC)。目前已知。 NK细胞发挥ADCC效应主要是通过其膜表面低亲和力FcγRⅢ(CD16)所介导的,IgG不仅起到连接靶细胞和效应细胞的作用,同时还刺激NK 细胞合成和分泌肿瘤坏死因子和γ干扰素等细胞因子,并释放颗粒,溶解靶细胞。嗜酸性粒细胞发挥ADCC作用是通过其FcεRⅡ和FcαR介导 的,嗜酸性粒细胞可脱颗粒释放碱性蛋白等,在杀伤寄生虫如蠕虫中发挥重要作用。 图2-10 抗体依赖的细胞介导的细胞毒作用(ADCC) 此外,人IgGFc段能非特异地与葡萄菌A蛋白(staphylococcus proteinA,SPA)结合,应用SPA可纯化IgG等抗体,或代替第二抗体用于标记 技术。 (四)通过胎盘 在人类,IgG是唯一可通过胎盘从母体转移给胎儿的Ig。IgG能选择性地与胎盘母体一侧的滋养层细胞结合,转移到滋养层细胞的吞饮泡 内,并主动外排到胎儿血循环中。IgG的这种功能与IgGFc片段结构有关,如切除Fc段后所剩余的Fab并不能通过胎盘。IgG通过胎盘的作用是