正在加载图片...
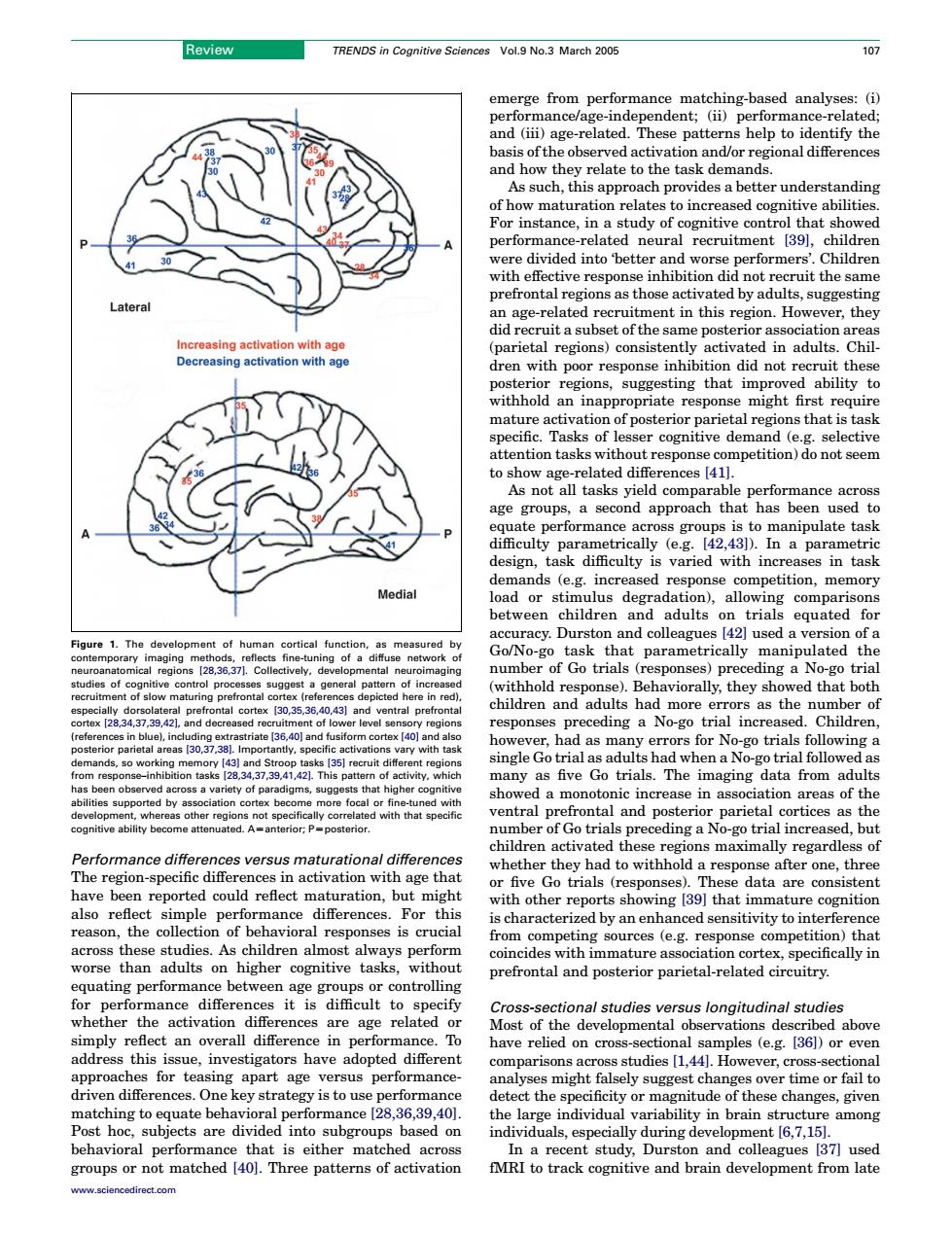
Review TRENDS in Cognitive Sciences Vol.9 No.3 March 2005 10> atchi and (iii)age-related As such,this approach provides a better underst anding performance-related neural recruitment 39,children prefrontal regions as those activated by adults. age-related ent in this region However,they reasing activatio arietal regions)consisten ctivated in adults Chil dren with p sugg ateng that impr mature activation of posterior parietal ecific. r cognitiv e demand lective show ag elated differences 41 As not app ametri var or ulus degradation),allowi ons dre on tria that the f Go trial sponses pre ding a no-g and adults had mo ra they as the pre ceding a No-go tria ine as five Go trials.The imaging data from adults mn asso ted withh a monotonic reas r of Go trial ding a no-go trial in d but en ance differer ces versus maturational differer ces n activation with age t n 11 five Go trials ing 39 that in ature cognitior this y an en eason the of avioral crucial e tiwitytointer cr lways pertorn ,specifically in prefrontal and posterior parietal-related circuitry het Cr ctional studies versus longitudinal studies Most of developmental observations T ed above this pted different udies (1.441.Ho or ever apar alyses might falsely sugges hanges over time or fail to 28,36,39,40 the ang ost hoc subj jects are divide into subgroups based on individuals.espe cially during development 6.7.15 matched study,Du collPerformance differences versus maturational differences The region-specific differences in activation with age that have been reported could reflect maturation, but might also reflect simple performance differences. For this reason, the collection of behavioral responses is crucial across these studies. As children almost always perform worse than adults on higher cognitive tasks, without equating performance between age groups or controlling for performance differences it is difficult to specify whether the activation differences are age related or simply reflect an overall difference in performance. To address this issue, investigators have adopted different approaches for teasing apart age versus performancedriven differences. One key strategy is to use performance matching to equate behavioral performance [28,36,39,40]. Post hoc, subjects are divided into subgroups based on behavioral performance that is either matched across groups or not matched [40]. Three patterns of activation emerge from performance matching-based analyses: (i) performance/age-independent; (ii) performance-related; and (iii) age-related. These patterns help to identify the basis of the observed activation and/or regional differences and how they relate to the task demands. As such, this approach provides a better understanding of how maturation relates to increased cognitive abilities. For instance, in a study of cognitive control that showed performance-related neural recruitment [39], children were divided into ‘better and worse performers’. Children with effective response inhibition did not recruit the same prefrontal regions as those activated by adults, suggesting an age-related recruitment in this region. However, they did recruit a subset of the same posterior association areas (parietal regions) consistently activated in adults. Children with poor response inhibition did not recruit these posterior regions, suggesting that improved ability to withhold an inappropriate response might first require mature activation of posterior parietal regions that is task specific. Tasks of lesser cognitive demand (e.g. selective attention tasks without response competition) do not seem to show age-related differences [41]. As not all tasks yield comparable performance across age groups, a second approach that has been used to equate performance across groups is to manipulate task difficulty parametrically (e.g. [42,43]). In a parametric design, task difficulty is varied with increases in task demands (e.g. increased response competition, memory load or stimulus degradation), allowing comparisons between children and adults on trials equated for accuracy. Durston and colleagues [42] used a version of a Go/No-go task that parametrically manipulated the number of Go trials (responses) preceding a No-go trial (withhold response). Behaviorally, they showed that both children and adults had more errors as the number of responses preceding a No-go trial increased. Children, however, had as many errors for No-go trials following a single Go trial as adults had when a No-go trial followed as many as five Go trials. The imaging data from adults showed a monotonic increase in association areas of the ventral prefrontal and posterior parietal cortices as the number of Go trials preceding a No-go trial increased, but children activated these regions maximally regardless of whether they had to withhold a response after one, three or five Go trials (responses). These data are consistent with other reports showing [39] that immature cognition is characterized by an enhanced sensitivity to interference from competing sources (e.g. response competition) that coincides with immature association cortex, specifically in prefrontal and posterior parietal-related circuitry. Cross-sectional studies versus longitudinal studies Most of the developmental observations described above have relied on cross-sectional samples (e.g. [36]) or even comparisons across studies [1,44]. However, cross-sectional analyses might falsely suggest changes over time or fail to detect the specificity or magnitude of these changes, given the large individual variability in brain structure among individuals, especially during development [6,7,15]. In a recent study, Durston and colleagues [37] used fMRI to track cognitive and brain development from late Figure 1. The development of human cortical function, as measured by contemporary imaging methods, reflects fine-tuning of a diffuse network of neuroanatomical regions [28,36,37]. Collectively, developmental neuroimaging studies of cognitive control processes suggest a general pattern of increased recruitment of slow maturing prefrontal cortex (references depicted here in red), especially dorsolateral prefrontal cortex [30,35,36,40,43] and ventral prefrontal cortex [28,34,37,39,42], and decreased recruitment of lower level sensory regions (references in blue), including extrastriate [36,40] and fusiform cortex [40] and also posterior parietal areas [30,37,38]. Importantly, specific activations vary with task demands, so working memory [43] and Stroop tasks [35] recruit different regions from response–inhibition tasks [28,34,37,39,41,42]. This pattern of activity, which has been observed across a variety of paradigms, suggests that higher cognitive abilities supported by association cortex become more focal or fine-tuned with development, whereas other regions not specifically correlated with that specific cognitive ability become attenuated. AZanterior; PZposterior. Review TRENDS in Cognitive Sciences Vol.9 No.3 March 2005 107 www.sciencedirect.com