正在加载图片...
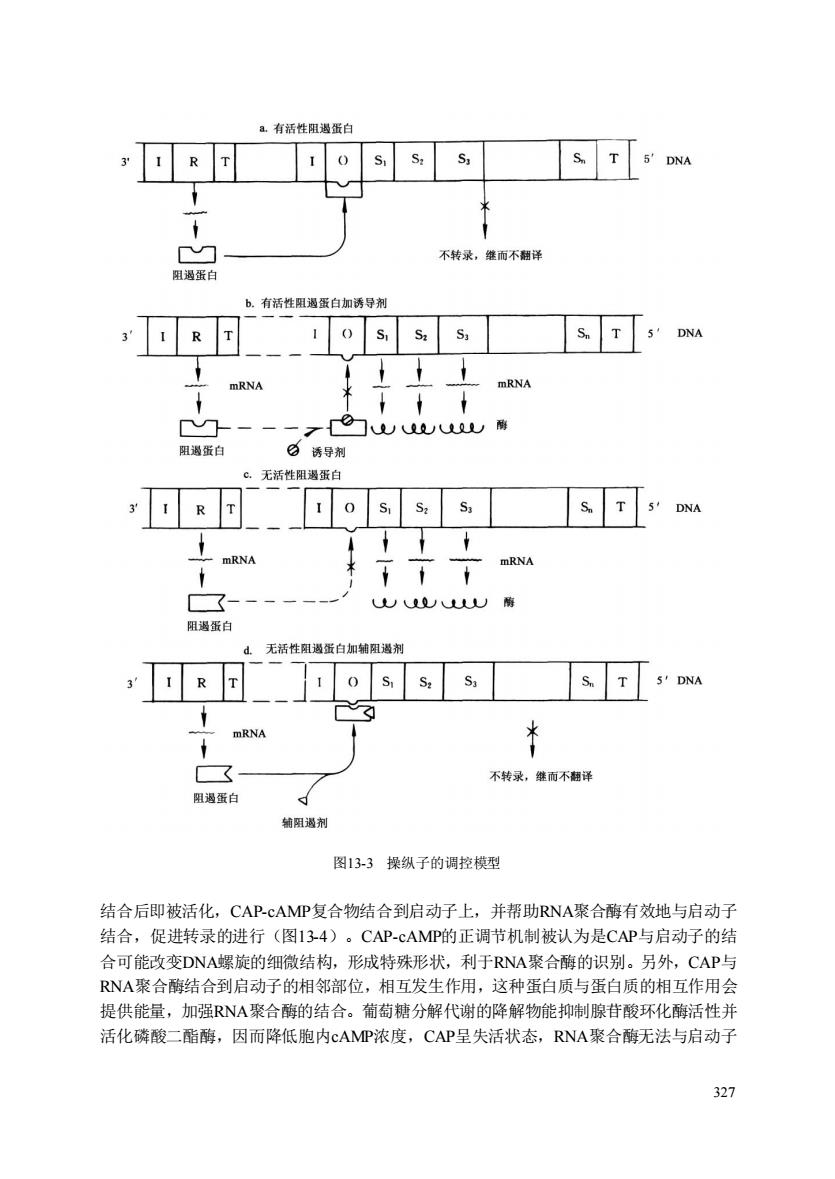
有括性阻渴蛋白 s 1 R T 1 o S.S.S, ST S'DNA M 不转来,继而不翻译 b.有活性阻遇蛋白加诱导剂 3'1 R T1 0 S,S.S,S.T 3 DA mRNA Z白刚 阻過蛋自 白诱导剂 。.无活性阻渴蛋白 日ss S.T s DNA A mRNA 了TfaM 一 UU南 阻遇蛋白 无活性蛋白如辅阻渴剂 ,I RT 1 o S.S.S, S.T 3'DNA ☒ mRNA * 不转录,雕而不相译 阳蛋白 菊阳遏剂 图13-3操纵子的调控模型 结合后即被活化,CAP-cAMP复合物结合到启动子上,并帮助RNA聚合酶有效地与启动子 结合,促进转录的进行(图134)。CAP-cAMP的正调节机制被认为是CAP与启动子的结 合可能改变DNA螺旋的细微结构,形城特殊形状,利于RNA聚合酶的识别。另外,CAP与 RNA聚合酶结合到启动子的相邻部位,相互发生作用,这种蛋白质与蛋白质的相互作用会 提供能量,加强RNA聚合酶的结合。葡萄糖分解代谢的降解物能抑制腺苷酸环化酶活性并 活化磷酸二酯酶,因而降低胞内cAMP浓度,CAP呈失活状态,RNA聚合酶无法与启动子 327 327 图13-3 操纵子的调控模型 结合后即被活化,CAP-cAMP复合物结合到启动子上,并帮助RNA聚合酶有效地与启动子 结合,促进转录的进行(图13-4)。CAP-cAMP的正调节机制被认为是CAP与启动子的结 合可能改变DNA螺旋的细微结构,形成特殊形状,利于RNA聚合酶的识别。另外,CAP与 RNA聚合酶结合到启动子的相邻部位,相互发生作用,这种蛋白质与蛋白质的相互作用会 提供能量,加强RNA聚合酶的结合。葡萄糖分解代谢的降解物能抑制腺苷酸环化酶活性并 活化磷酸二酯酶,因而降低胞内cAMP浓度,CAP呈失活状态,RNA聚合酶无法与启动子