正在加载图片...
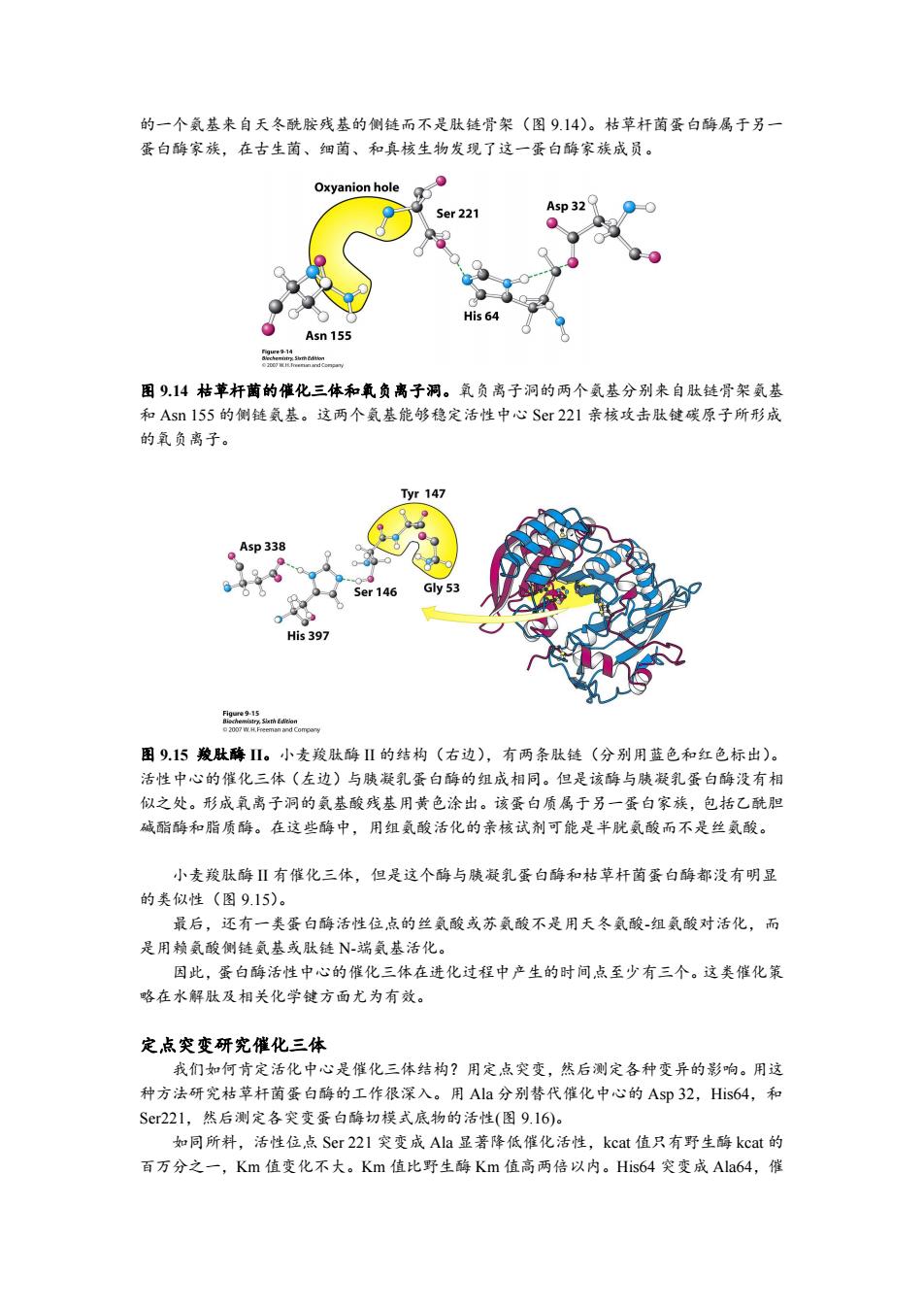
的一个氨基来自天冬酰胺残基的侧链而不是肽链骨架(图9.14)。枯草杆菌蛋白酶属于另一 蛋白酶家族,在古生菌、细菌、和真核生物发现了这一蛋白酶家族成员。 Oxyanion hole Ser 221 Asp 32 His 64 Asn 155 图914枯苹杆菌的催化三体和氧负离子洞。氧负离子洞的两个气基分别来自肽链骨架氨基 和Asn155的侧链氦基。这两个氨基能够稳定活性中心Sr221亲核攻击肽键碳原子所形成 的氧负离子。 yr147 Asp 338 Ser 146 Gly 53 0 His 397 feca5shEon 图9.15羧肽酶Ⅱ。小麦羧肽酶Ⅱ的结构(右边),有两条肽链(分别用蓝色和红色标出)。 活性中心的催化三体(左边)与胰凝乳蛋白酶的组成相同。但是该酶与胰凝乳蛋白酶没有相 似之处。形成氧离子洞的氨基酸残基用黄色涂出。该蛋白质属于另一蛋白家族,包括乙酰胆 碱酯酶和脂质酶。在这些酶中,用组气酸活化的亲核试剂可能是半胱氨酸而不是丝氨酸。 小麦羧肽酶Ⅱ有催化三体,但是这个酶与胰凝乳蛋白酶和枯草杆菌蛋白酶都没有明显 的类似性(图9.15)。 最后,还有一类蛋白酶活性位点的丝氨酸或苏氨酸不是用天冬氨酸-组氨酸对活化,而 是用赖氨酸侧链氨基或肽链N-瑞氦基活化。 因此,蛋白酶活性中心的催化三体在进化过程中产生的时间点至少有三个。这类催化策 略在水解肽及相关化学键方面尤为有效。 定,点突变研究催化三体 我们如何肯定活化中心是催化三体结构?用定点突变,然后测定各种变异的影响。用这 种方法研究枯草杆菌蛋白酶的工作很深入。用Ala分别替代催化中心的Asp32,His64,和 Sr221,然后测定各突变蛋白酶切模式底物的活性(图9.16)。 如同所料,活性位点Ser221突变成Ala显著降低催化活性,kcat值只有野生酶kcat的 百万分之一,Km值变化不大。Km值比野生酶Km值高两倍以内。His64突变成Ala64,催的一个氨基来自天冬酰胺残基的侧链而不是肽链骨架(图 9.14)。枯草杆菌蛋白酶属于另一 蛋白酶家族,在古生菌、细菌、和真核生物发现了这一蛋白酶家族成员。 图 9.14 枯草杆菌的催化三体和氧负离子洞。氧负离子洞的两个氨基分别来自肽链骨架氨基 和 Asn 155 的侧链氨基。这两个氨基能够稳定活性中心 Ser 221 亲核攻击肽键碳原子所形成 的氧负离子。 图 9.15 羧肽酶 II。小麦羧肽酶 II 的结构(右边),有两条肽链(分别用蓝色和红色标出)。 活性中心的催化三体(左边)与胰凝乳蛋白酶的组成相同。但是该酶与胰凝乳蛋白酶没有相 似之处。形成氧离子洞的氨基酸残基用黄色涂出。该蛋白质属于另一蛋白家族,包括乙酰胆 碱酯酶和脂质酶。在这些酶中,用组氨酸活化的亲核试剂可能是半胱氨酸而不是丝氨酸。 小麦羧肽酶 II 有催化三体,但是这个酶与胰凝乳蛋白酶和枯草杆菌蛋白酶都没有明显 的类似性(图 9.15)。 最后,还有一类蛋白酶活性位点的丝氨酸或苏氨酸不是用天冬氨酸-组氨酸对活化,而 是用赖氨酸侧链氨基或肽链 N-端氨基活化。 因此,蛋白酶活性中心的催化三体在进化过程中产生的时间点至少有三个。这类催化策 略在水解肽及相关化学键方面尤为有效。 定点突变研究催化三体 我们如何肯定活化中心是催化三体结构?用定点突变,然后测定各种变异的影响。用这 种方法研究枯草杆菌蛋白酶的工作很深入。用 Ala 分别替代催化中心的 Asp 32,His64,和 Ser221,然后测定各突变蛋白酶切模式底物的活性(图 9.16)。 如同所料,活性位点 Ser 221 突变成 Ala 显著降低催化活性,kcat 值只有野生酶 kcat 的 百万分之一,Km 值变化不大。Km 值比野生酶 Km 值高两倍以内。His64 突变成 Ala64,催