正在加载图片...
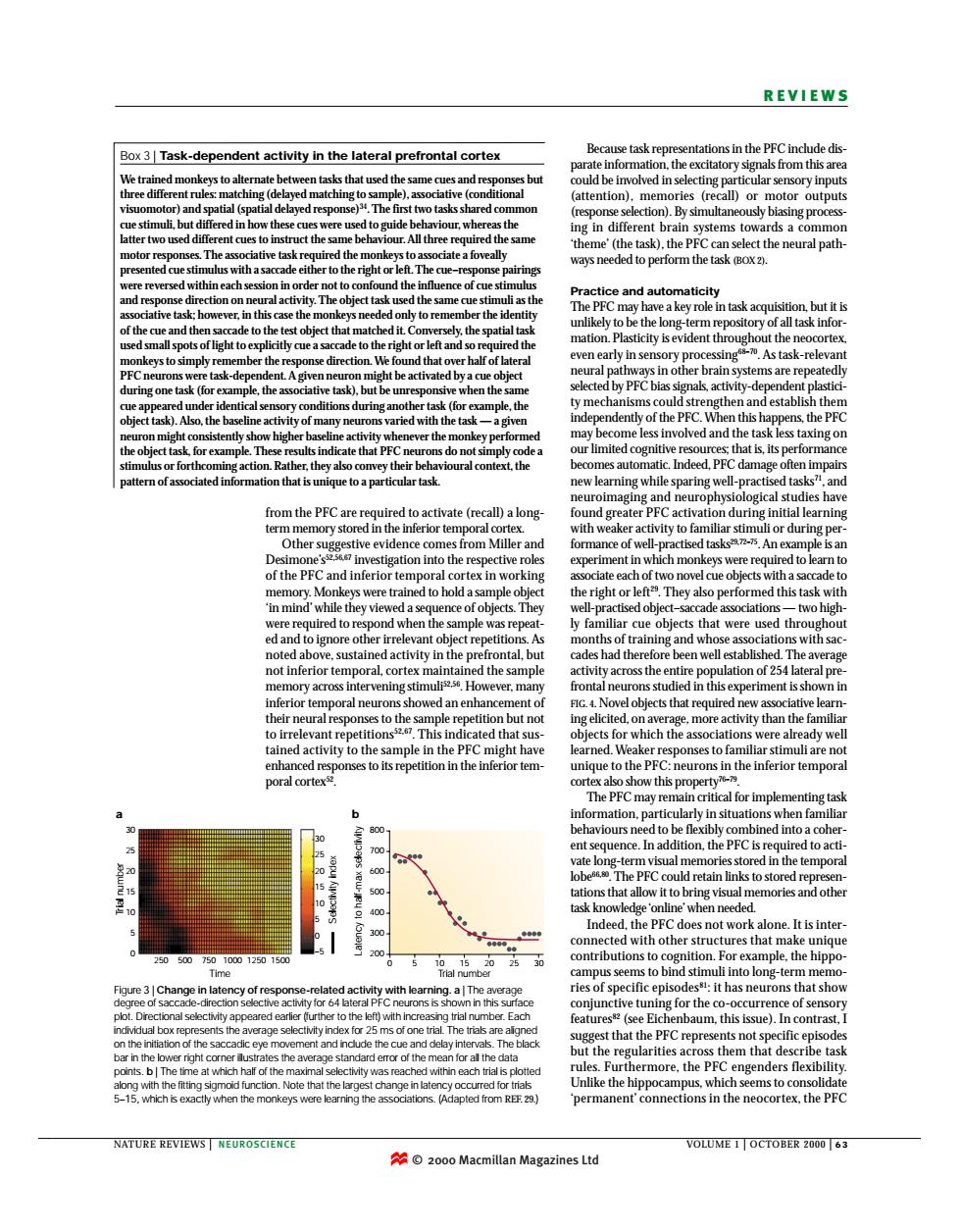
REVIEWS Box3Task-dependent activity in the lateral prefrontal cortex ons in the PFC inchde d Ve tra ed res iel.asodatiee atention,mmopyiSf ne PFC avs n her to the right or】 e object tobe th de to the ry pro by PFCbia ny of the PFC. en this h that is.its ida nes from Miller ex in w rep me s tha nedactiytotheanmpleinheECtg ed.Weak poral cortex with othe 05 for the) that the PFC repr s not so mean for al rthermore.the PFC y occu ermanent' 2000 Macmillan Magazines Ltofrom the PFC are required to activate (recall) a longterm memory stored in the inferior temporal cortex. Other suggestive evidence comes from Miller and Desimone’s52,56,67 investigation into the respective roles of the PFC and inferior temporal cortex in working memory. Monkeys were trained to hold a sample object ‘in mind’ while they viewed a sequence of objects. They were required to respond when the sample was repeated and to ignore other irrelevant object repetitions. As noted above, sustained activity in the prefrontal, but not inferior temporal, cortex maintained the sample memory across intervening stimuli52,56. However, many inferior temporal neurons showed an enhancement of their neural responses to the sample repetition but not to irrelevant repetitions52,67. This indicated that sustained activity to the sample in the PFC might have enhanced responses to its repetition in the inferior temporal cortex52. Box 3 | Task-dependent activity in the lateral prefrontal cortex We trained monkeys to alternate between tasks that used the same cues and responses but three different rules: matching (delayed matching to sample), associative (conditional visuomotor) and spatial (spatial delayed response)34. The first two tasks shared common cue stimuli, but differed in how these cues were used to guide behaviour, whereas the latter two used different cues to instruct the same behaviour. All three required the same motor responses. The associative task required the monkeys to associate a foveally presented cue stimulus with a saccade either to the right or left. The cue–response pairings were reversed within each session in order not to confound the influence of cue stimulus and response direction on neural activity. The object task used the same cue stimuli as the associative task; however, in this case the monkeys needed only to remember the identity of the cue and then saccade to the test object that matched it. Conversely, the spatial task used small spots of light to explicitly cue a saccade to the right or left and so required the monkeys to simply remember the response direction. We found that over half of lateral PFC neurons were task-dependent. A given neuron might be activated by a cue object during one task (for example, the associative task), but be unresponsive when the same cue appeared under identical sensory conditions during another task (for example, the object task). Also, the baseline activity of many neurons varied with the task — a given neuron might consistently show higher baseline activity whenever the monkey performed the object task, for example. These results indicate that PFC neurons do not simply code a stimulus or forthcoming action. Rather, they also convey their behavioural context, the pattern of associated information that is unique to a particular task. NATURE REVIEWS | NEUROSCIENCE VOLUME 1 | OCTOBER 2000 | 6 3 REVIEWS Because task representations in the PFC include disparate information, the excitatory signals from this area could be involved in selecting particular sensory inputs (attention), memories (recall) or motor outputs (response selection). By simultaneously biasing processing in different brain systems towards a common ‘theme’ (the task), the PFC can select the neural pathways needed to perform the task (BOX 2). Practice and automaticity The PFC may have a key role in task acquisition, but it is unlikely to be the long-term repository of all task information. Plasticity is evident throughout the neocortex, even early in sensory processing68–70. As task-relevant neural pathways in other brain systems are repeatedly selected by PFC bias signals, activity-dependent plasticity mechanisms could strengthen and establish them independently of the PFC. When this happens, the PFC may become less involved and the task less taxing on our limited cognitive resources; that is, its performance becomes automatic. Indeed, PFC damage often impairs new learning while sparing well-practised tasks71, and neuroimaging and neurophysiological studies have found greater PFC activation during initial learning with weaker activity to familiar stimuli or during performance of well-practised tasks29,72–75. An example is an experiment in which monkeys were required to learn to associate each of two novel cue objects with a saccade to the right or left29. They also performed this task with well-practised object–saccade associations — two highly familiar cue objects that were used throughout months of training and whose associations with saccades had therefore been well established. The average activity across the entire population of 254 lateral prefrontal neurons studied in this experiment is shown in FIG. 4. Novel objects that required new associative learning elicited, on average, more activity than the familiar objects for which the associations were already well learned. Weaker responses to familiar stimuli are not unique to the PFC: neurons in the inferior temporal cortex also show this property76–79. The PFC may remain critical for implementing task information, particularly in situations when familiar behaviours need to be flexibly combined into a coherent sequence. In addition, the PFC is required to activate long-term visual memories stored in the temporal lobe66,80. The PFC could retain links to stored representations that allow it to bring visual memories and other task knowledge ‘online’ when needed. Indeed, the PFC does not work alone. It is interconnected with other structures that make unique contributions to cognition. For example, the hippocampus seems to bind stimuli into long-term memories of specific episodes81: it has neurons that show conjunctive tuning for the co-occurrence of sensory features82 (see Eichenbaum, this issue). In contrast, I suggest that the PFC represents not specific episodes but the regularities across them that describe task rules. Furthermore, the PFC engenders flexibility. Unlike the hippocampus, which seems to consolidate ‘permanent’ connections in the neocortex, the PFC 30 800 a b 25 10 5 20 15 0 250 500 750 1000 1250 1500 30 20 25 5 0 15 10 –5 700 600 500 400 300 200 0 5 Latency to half-max selectivity Trial number Selectivity index 10 15 Time Trial number 20 25 30 Figure 3 | Change in latency of response-related activity with learning. a | The average degree of saccade-direction selective activity for 64 lateral PFC neurons is shown in this surface plot. Directional selectivity appeared earlier (further to the left) with increasing trial number. Each individual box represents the average selectivity index for 25 ms of one trial. The trials are aligned on the initiation of the saccadic eye movement and include the cue and delay intervals. The black bar in the lower right corner illustrates the average standard error of the mean for all the data points. b | The time at which half of the maximal selectivity was reached within each trial is plotted along with the fitting sigmoid function. Note that the largest change in latency occurred for trials 5–15, which is exactly when the monkeys were learning the associations. (Adapted from REF. 29.) © 2000 Macmillan Magazines Ltd