正在加载图片...
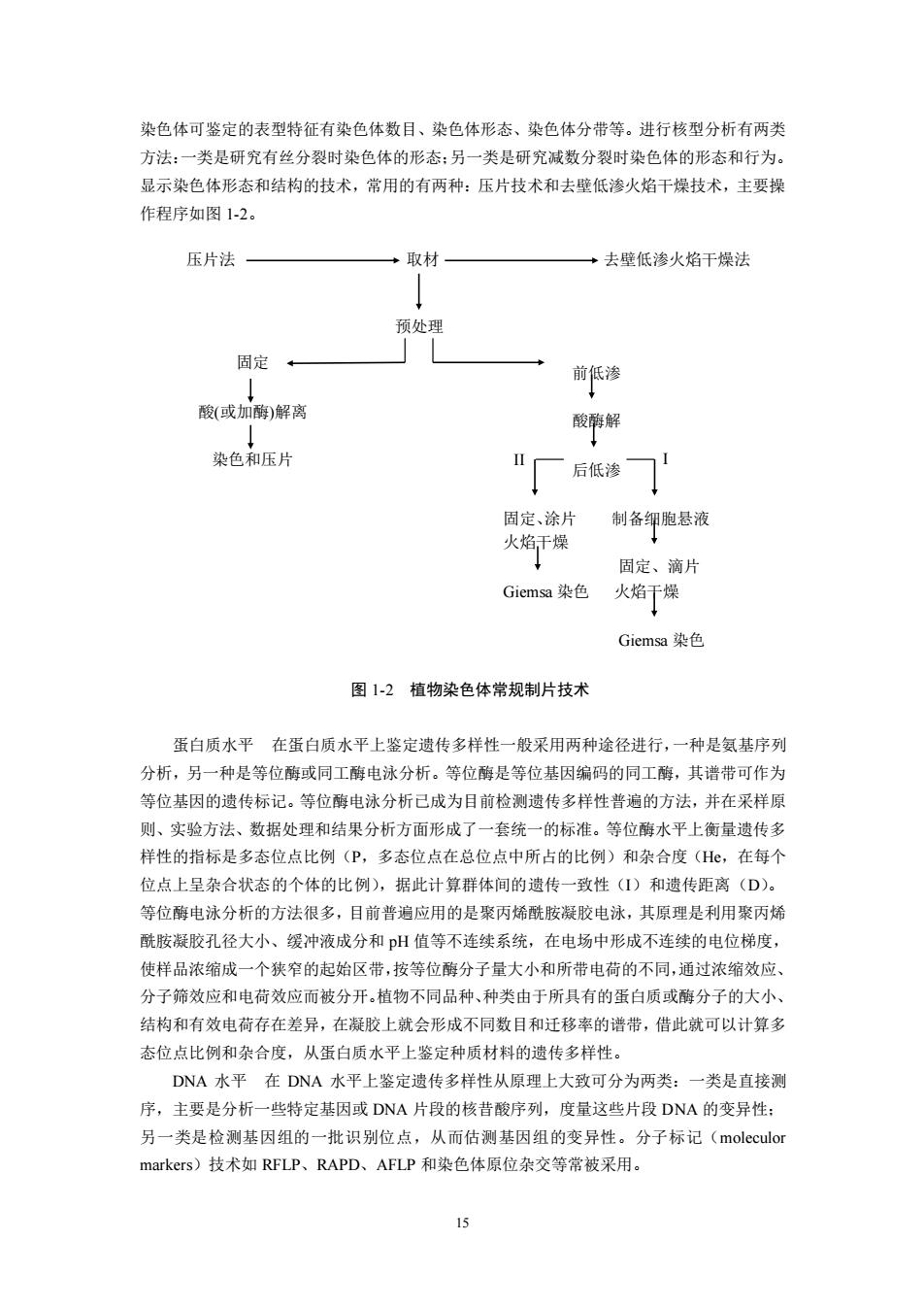
染色体可鉴定的表型特征有染色体数目、染色体形态、染色体分带等。进行核型分析有两类 方法:一类是研究有丝分裂时染色体的形态:另一类是研究减数分裂时染色体的形态和行为。 显示染色体形态和结构的技术,常用的有两种:压片技术和去壁低渗火焰干燥技术,主要操 作程序如图1-2。 压片法 取材 ·去壁低渗火焰干燥法 预处理 固定 前低渗 ↓ 酸(或加酶)解离 酸酶解 染色和压片 "厂 后低渗 固定、涂片 制备细胞悬液 火焰干燥 固定、滴片 Giemsa染色 火焰干燥 Giemsa染色 图1-2植物染色体常规制片技术 蛋白质水平在蛋白质水平上鉴定遗传多样性一般采用两种途径进行,一种是氨基序列 分析,另一种是等位酶或同工酶电泳分析。等位酶是等位基因编码的同工酶,其谱带可作为 等位基因的遗传标记。等位酶电泳分析己成为目前检测遗传多样性普遍的方法,并在采样原 则、实验方法、数据处理和结果分析方面形成了一套统一的标准。等位酶水平上衡量遗传多 样性的指标是多态位点比例(P,多态位点在总位点中所占的比例)和杂合度(H,在每个 位点上呈杂合状态的个体的比例),据此计算群体间的遗传一致性(I)和遗传距离(D)。 等位酶电泳分析的方法很多,目前普遍应用的是聚丙烯酰胺凝胶电泳,其原理是利用聚丙烯 酰胺凝胶孔径大小、缓冲液成分和pH值等不连续系统,在电场中形成不连续的电位梯度, 使样品浓缩成一个狭窄的起始区带,按等位酶分子量大小和所带电荷的不同,通过浓缩效应、 分子筛效应和电荷效应而被分开。植物不同品种、种类由于所具有的蛋白质或酶分子的大小、 结构和有效电荷存在差异,在凝胶上就会形成不同数目和迁移率的谱带,借此就可以计算多 态位点比例和杂合度,从蛋白质水平上鉴定种质材料的遗传多样性。 DNA水平在DNA水平上鉴定遗传多样性从原理上大致可分为两类:一类是直接测 序,主要是分析一些特定基因或DNA片段的核昔酸序列,度量这些片段DNA的变异性: 另一类是检测基因组的一批识别位点,从而估测基因组的变异性。分子标记(moleculor markers)技术如RFLP、RAPD、AFLP和染色体原位杂交等常被采用。 公15 染色体可鉴定的表型特征有染色体数目、染色体形态、染色体分带等。进行核型分析有两类 方法:一类是研究有丝分裂时染色体的形态;另一类是研究减数分裂时染色体的形态和行为。 显示染色体形态和结构的技术,常用的有两种:压片技术和去壁低渗火焰干燥技术,主要操 作程序如图 1-2。 图 1-2 植物染色体常规制片技术 蛋白质水平 在蛋白质水平上鉴定遗传多样性一般采用两种途径进行,一种是氨基序列 分析,另一种是等位酶或同工酶电泳分析。等位酶是等位基因编码的同工酶,其谱带可作为 等位基因的遗传标记。等位酶电泳分析已成为目前检测遗传多样性普遍的方法,并在采样原 则、实验方法、数据处理和结果分析方面形成了一套统一的标准。等位酶水平上衡量遗传多 样性的指标是多态位点比例(P,多态位点在总位点中所占的比例)和杂合度(He,在每个 位点上呈杂合状态的个体的比例),据此计算群体间的遗传一致性(I)和遗传距离(D)。 等位酶电泳分析的方法很多,目前普遍应用的是聚丙烯酰胺凝胶电泳,其原理是利用聚丙烯 酰胺凝胶孔径大小、缓冲液成分和 pH 值等不连续系统,在电场中形成不连续的电位梯度, 使样品浓缩成一个狭窄的起始区带,按等位酶分子量大小和所带电荷的不同,通过浓缩效应、 分子筛效应和电荷效应而被分开。植物不同品种、种类由于所具有的蛋白质或酶分子的大小、 结构和有效电荷存在差异,在凝胶上就会形成不同数目和迁移率的谱带,借此就可以计算多 态位点比例和杂合度,从蛋白质水平上鉴定种质材料的遗传多样性。 DNA 水平 在 DNA 水平上鉴定遗传多样性从原理上大致可分为两类:一类是直接测 序,主要是分析一些特定基因或 DNA 片段的核昔酸序列,度量这些片段 DNA 的变异性; 另一类是检测基因组的一批识别位点,从而估测基因组的变异性。分子标记(moleculor markers)技术如 RFLP、RAPD、AFLP 和染色体原位杂交等常被采用。 压片法 取材 去壁低渗火焰干燥法 固定 酸(或加酶)解离 染色和压片 预处理 前低渗 酸酶解 后低渗 固定、涂片 制备细胞悬液 火焰干燥 固定、滴片 Giemsa 染色 火焰干燥 Giemsa 染色 II I