正在加载图片...
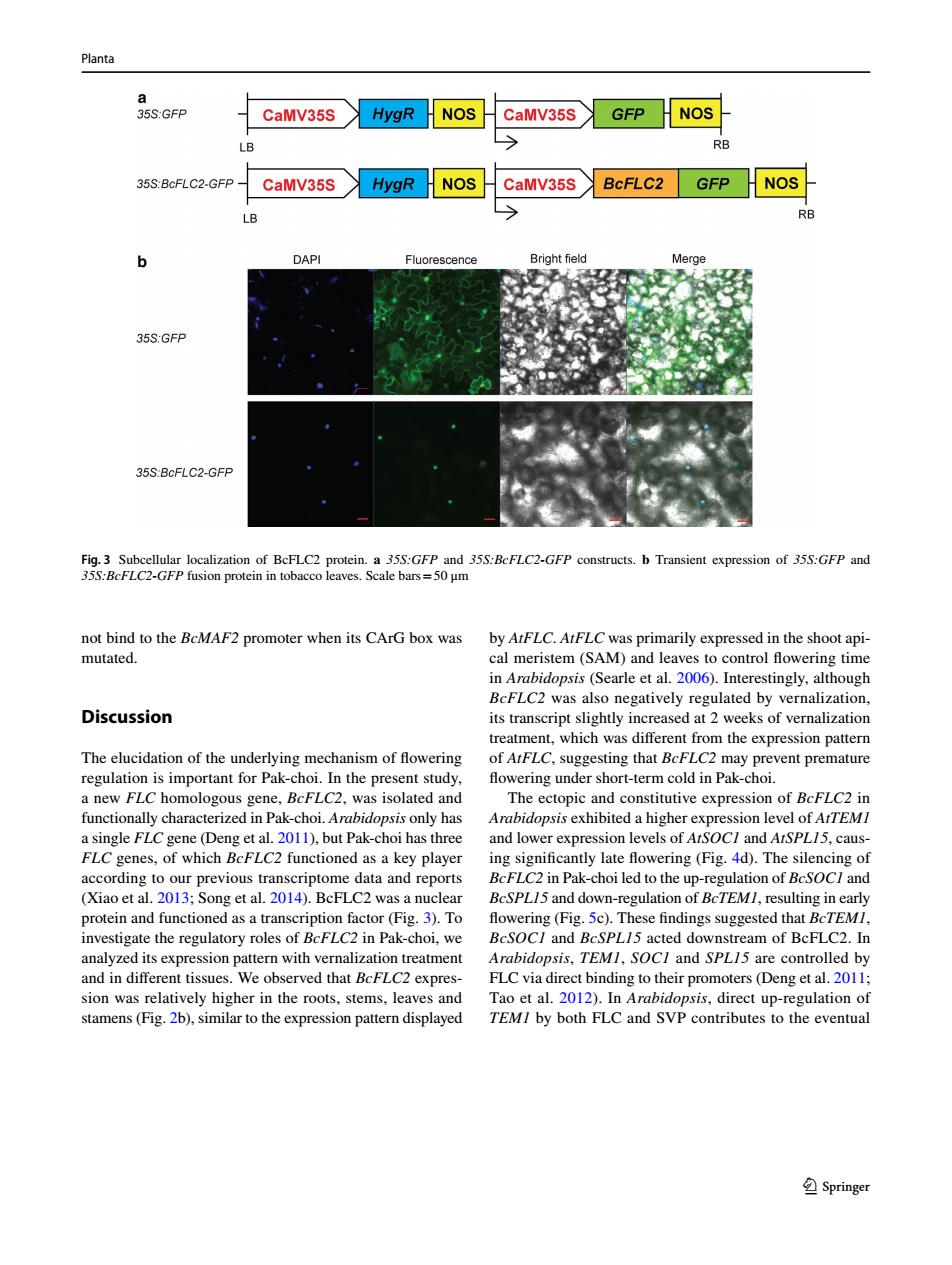
Planta a 35S:GFP CaMV35S HygR NOS CaMV35S GFP NOS LB RB 35S:BcFLC2-GFP CaMV35S HygR NOS CaMV35S BcFLC2 GFP HNOS LB RB b DAPI Fluorescence Bright field Merge 35S:GFP 35S:BcFLC2-GFP Fig.3 Subcellular localization of BcFLC2 protein.a 35S:GFP and 35S:BcFLC2-GFP constructs.b Transient expression of 35S:GFP and 35S:BcFLC2-GFP fusion protein in tobacco leaves.Scale bars=50 um not bind to the BcMAF2 promoter when its CArG box was by AtFLC.AtFLC was primarily expressed in the shoot api- mutated. cal meristem (SAM)and leaves to control flowering time in Arabidopsis(Searle et al.2006).Interestingly,although BcFLC2 was also negatively regulated by vernalization, Discussion its transcript slightly increased at 2 weeks of vernalization treatment,which was different from the expression pattern The elucidation of the underlying mechanism of flowering of AtFLC,suggesting that BcFLC2 may prevent premature regulation is important for Pak-choi.In the present study, flowering under short-term cold in Pak-choi. a new FLC homologous gene,BcFLC2,was isolated and The ectopic and constitutive expression of BcFLC2 in functionally characterized in Pak-choi.Arabidopsis only has Arabidopsis exhibited a higher expression level of AtTEMI a single FLC gene(Deng et al.2011),but Pak-choi has three and lower expression levels of AtSOC/and AtSPL15,caus- FLC genes,of which BcFLC2 functioned as a key player ing significantly late flowering (Fig.4d).The silencing of according to our previous transcriptome data and reports BcFLC2 in Pak-choi led to the up-regulation of BcSOCI and (Xiao et al.2013;Song et al.2014).BcFLC2 was a nuclear BcSPLI5 and down-regulation of BcTEMI,resulting in early protein and functioned as a transcription factor(Fig.3).To flowering(Fig.5c).These findings suggested that BcTEMI, investigate the regulatory roles of BcFLC2 in Pak-choi,we BcSOC/and BcSPL/5 acted downstream of BcFLC2.In analyzed its expression pattern with vernalization treatment Arabidopsis,TEMI,SOCI and SPL15 are controlled by and in different tissues.We observed that BcFLC2 expres- FLC via direct binding to their promoters (Deng et al.2011; sion was relatively higher in the roots,stems,leaves and Tao et al.2012).In Arabidopsis,direct up-regulation of stamens(Fig.2b),similar to the expression pattern displayed TEM/by both FLC and SVP contributes to the eventual SpringerPlanta 1 3 not bind to the BcMAF2 promoter when its CArG box was mutated. Discussion The elucidation of the underlying mechanism of fowering regulation is important for Pak-choi. In the present study, a new FLC homologous gene, BcFLC2, was isolated and functionally characterized in Pak-choi. Arabidopsis only has a single FLC gene (Deng et al. 2011), but Pak-choi has three FLC genes, of which BcFLC2 functioned as a key player according to our previous transcriptome data and reports (Xiao et al. 2013; Song et al. 2014). BcFLC2 was a nuclear protein and functioned as a transcription factor (Fig. 3). To investigate the regulatory roles of BcFLC2 in Pak-choi, we analyzed its expression pattern with vernalization treatment and in diferent tissues. We observed that BcFLC2 expression was relatively higher in the roots, stems, leaves and stamens (Fig. 2b), similar to the expression pattern displayed by AtFLC. AtFLC was primarily expressed in the shoot apical meristem (SAM) and leaves to control fowering time in Arabidopsis (Searle et al. 2006). Interestingly, although BcFLC2 was also negatively regulated by vernalization, its transcript slightly increased at 2 weeks of vernalization treatment, which was diferent from the expression pattern of AtFLC, suggesting that BcFLC2 may prevent premature fowering under short-term cold in Pak-choi. The ectopic and constitutive expression of BcFLC2 in Arabidopsis exhibited a higher expression level of AtTEM1 and lower expression levels of AtSOC1 and AtSPL15, causing signifcantly late fowering (Fig. 4d). The silencing of BcFLC2 in Pak-choi led to the up-regulation of BcSOC1 and BcSPL15 and down-regulation of BcTEM1, resulting in early fowering (Fig. 5c). These fndings suggested that BcTEM1, BcSOC1 and BcSPL15 acted downstream of BcFLC2. In Arabidopsis, TEM1, SOC1 and SPL15 are controlled by FLC via direct binding to their promoters (Deng et al. 2011; Tao et al. 2012). In Arabidopsis, direct up-regulation of TEM1 by both FLC and SVP contributes to the eventual Fig. 3 Subcellular localization of BcFLC2 protein. a 35S:GFP and 35S:BcFLC2-GFP constructs. b Transient expression of 35S:GFP and 35S:BcFLC2-GFP fusion protein in tobacco leaves. Scale bars=50 μm