正在加载图片...
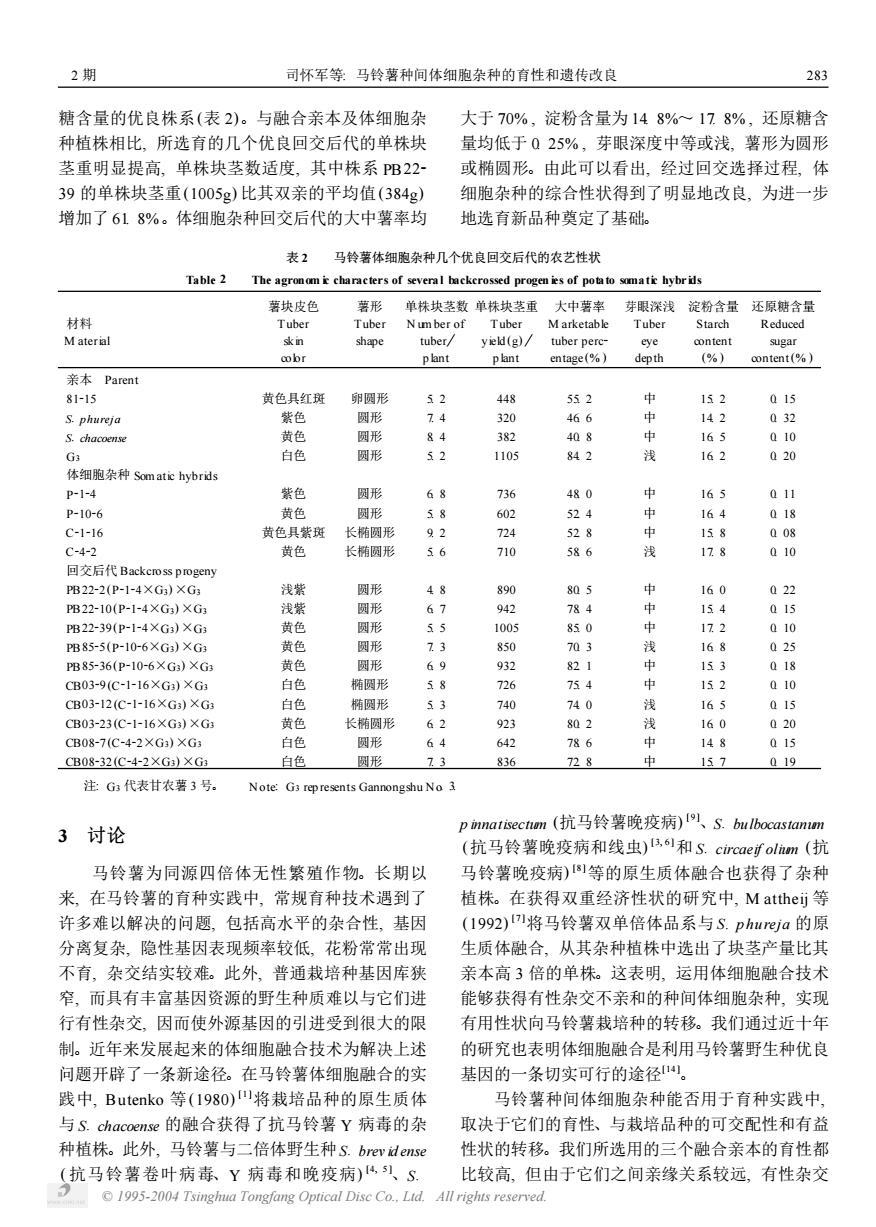
2期 司怀军等:马铃薯种间体细胞杂种的育性和遗传改良 283 糖含量的优良株系(表2)。与融合亲本及体细胞杂 大于70%,淀粉含量为148%~178%,还原糖含 种植株相比,所选育的几个优良回交后代的单株块 量均低于Q25%,芽眼深度中等或浅,薯形为圆形 茎重明显提高,单株块茎数适度,其中株系PB22 或椭圆形。由此可以看出,经过回交选择过程,体 39的单株块茎重(1005g)比其双亲的平均值(384g) 细胞杂种的综合性状得到了明显地改良,为进一步 增加了618%。体细胞杂种回交后代的大中薯率均 地选育新品种奠定了基础。 表2 马铃薯体细胞杂种几个优良回交后代的农艺性状 Table 2 The agronom i characters of several backerossed progen ies of potato somatic hybrids 薯块皮色 薯形 单株块茎数单株块茎重大中薯率 芽眼深浅淀粉含量还原糖含量 材料 Tuber Tuber Num ber of Tuber M arketable Tuber Starch Reduced M aterial skin shape tuber/ yield(g)/ tuber perc- eye content sugar cobr plant plant entage(%) depth (%) content(%) 亲本Parent 81-15 黄色具红斑 卵圆形 52 448 552 中 152 015 S.phureja 紫色 圆形 74 320 466 中 142 032 S.chacoense 黄色 圆形 84 382 408 中 165 010 G3 白色 圆形 52 1105 842 浅 162 020 体细胞杂种Somatic hybrids p-1-4 紫色 圆形 68 736 480 中 165 011 P-10-6 黄色 圆形 58 602 524 中 164 018 C-1-16 黄色具紫斑 长椭圆形 92 724 528 中 158 008 C-4-2 黄色 长椭圆形 56 710 586 浅 178 010 回交后代Backcross progeny PB22-2(P-1-4×G)×G 浅紫 圆形 48 890 805 必 160 022 PB22-10(P-1-4×G3)×G3 浅紫 圆形 67 942 784 中 154 015 PB22-39(p-1-4×G3)×G3 黄色 圆形 55 1005 850 中 172 010 PB85-5(P-10-6XG3)×G3 黄色 圆形 13 850 703 浅 168 025 PB85-36(P-10-6×G)×G 黄色 圆形 69 932 821 中 153 018 CB03-9(C-1-16XG)×G3 白色 椭圆形 58 726 754 中 152 010 CB03-12(C-1-16XG3)XG 白色 椭圆形 53 740 740 浅 165 015 CB03-23(C-1-16×G3)×G 黄色 长椭圆形 62 923 802 浅 160 020 CB08-7(C-4-2×G3)×G3 白色 圆形 64 642 786 中 148 015 CB08-32(C-4-2XG)×G3 白色 圆形 73 836 728 中 157 019 注G代表甘农薯3号。 Note:G3 represents Gannongshu No 3 3讨论 pinnatisectmn(抗马铃薯晚疫病)1、S.bulbocastamm (抗马铃薯晚疫病和线虫)B1和S.circaeifolim(抗 马铃薯为同源四倍体无性繁殖作物。长期以 马铃薯晚疫病)]等的原生质体融合也获得了杂种 来,在马铃薯的育种实践中,常规育种技术遇到了 植株。在获得双重经济性状的研究中,M attheij等 许多难以解决的问题,包括高水平的杂合性,基因 (1992)I将马铃薯双单倍体品系与S.phureja的原 分离复杂,隐性基因表现频率较低,花粉常常出现 生质体融合,从其杂种植株中选出了块茎产量比其 不育,杂交结实较难。此外,普通栽培种基因库狭 亲本高3倍的单株。这表明,运用体细胞融合技术 窄,而具有丰富基因资源的野生种质难以与它们进 能够获得有性杂交不亲和的种间体细胞杂种,实现 行有性杂交,因而使外源基因的引进受到很大的限 有用性状向马铃薯栽培种的转移。我们通过近十年 制。近年来发展起来的体细胞融合技术为解决上述 的研究也表明体细胞融合是利用马铃薯野生种优良 问题开辟了一条新途径。在马铃薯体细胞融合的实 基因的一条切实可行的途径。 践中,Butenko等(1980))将栽培品种的原生质体 马铃薯种间体细胞杂种能否用于育种实践中, 与S.chacoense的融合获得了抗马铃薯Y病毒的杂 取决于它们的育性、与栽培品种的可交配性和有益 种植株。此外,马铃薯与二倍体野生种S.brevidense 性状的转移。我们所选用的三个融合亲本的育性都 (抗马铃薯卷叶病毒、Y病毒和晚疫病)、S. 比较高,但由于它们之间亲缘关系较远,有性杂交 C 1995-2004 Tsinghua Tongfang Optical Disc Co..Ltd.All rights reserved.糖含量的优良株系(表 2)。与融合亲本及体细胞杂 种植株相比, 所选育的几个优良回交后代的单株块 茎重明显提高, 单株块茎数适度, 其中株系 PB222 39 的单株块茎重(1005g) 比其双亲的平均值(384g) 增加了 61. 8%。体细胞杂种回交后代的大中薯率均 大于 70% , 淀粉含量为 14. 8%~ 17. 8% , 还原糖含 量均低于 0. 25% , 芽眼深度中等或浅, 薯形为圆形 或椭圆形。由此可以看出, 经过回交选择过程, 体 细胞杂种的综合性状得到了明显地改良, 为进一步 地选育新品种奠定了基础。 表 2 马铃薯体细胞杂种几个优良回交后代的农艺性状 Table 2 The agronom ic characters of several backcrossed progen ies of potato somatic hybr ids 材料 M aterial 薯块皮色 Tuber skin color 薯形 Tuber shape 单株块茎数 N um ber of tuberö p lant 单株块茎重 Tuber yield (g)ö p lant 大中薯率 M arketable tuber perc2 entage (% ) 芽眼深浅 Tuber eye dep th 淀粉含量 Starch content (% ) 还原糖含量 Reduced sugar content(% ) 亲本 Parent 81215 黄色具红斑 卵圆形 5. 2 448 55. 2 中 15. 2 0. 15 S. phureja 紫色 圆形 7. 4 320 46. 6 中 14. 2 0. 32 S. chacoense 黄色 圆形 8. 4 382 40. 8 中 16. 5 0. 10 G3 白色 圆形 5. 2 1105 84. 2 浅 16. 2 0. 20 体细胞杂种 Som atic hybrids P2124 紫色 圆形 6. 8 736 48. 0 中 16. 5 0. 11 P21026 黄色 圆形 5. 8 602 52. 4 中 16. 4 0. 18 C21216 黄色具紫斑 长椭圆形 9. 2 724 52. 8 中 15. 8 0. 08 C2422 黄色 长椭圆形 5. 6 710 58. 6 浅 17. 8 0. 10 回交后代 Backcross p rogeny PB2222 (P2124×G3) ×G3 浅紫 圆形 4. 8 890 80. 5 中 16. 0 0. 22 PB22210 (P2124×G3) ×G3 浅紫 圆形 6. 7 942 78. 4 中 15. 4 0. 15 PB22239 (P2124×G3) ×G3 黄色 圆形 5. 5 1005 85. 0 中 17. 2 0. 10 PB8525 (P21026×G3) ×G3 黄色 圆形 7. 3 850 70. 3 浅 16. 8 0. 25 PB85236 (P21026×G3) ×G3 黄色 圆形 6. 9 932 82. 1 中 15. 3 0. 18 CB0329 (C21216×G3) ×G3 白色 椭圆形 5. 8 726 75. 4 中 15. 2 0. 10 CB03212 (C21216×G3) ×G3 白色 椭圆形 5. 3 740 74. 0 浅 16. 5 0. 15 CB03223 (C21216×G3) ×G3 黄色 长椭圆形 6. 2 923 80. 2 浅 16. 0 0. 20 CB0827 (C2422×G3) ×G3 白色 圆形 6. 4 642 78. 6 中 14. 8 0. 15 CB08232 (C2422×G3) ×G3 白色 圆形 7. 3 836 72. 8 中 15. 7 0. 19 注: G3 代表甘农薯 3 号。 Note: G3 rep resents Gannongshu No. 3. 3 讨论 马铃薯为同源四倍体无性繁殖作物。长期以 来, 在马铃薯的育种实践中, 常规育种技术遇到了 许多难以解决的问题, 包括高水平的杂合性, 基因 分离复杂, 隐性基因表现频率较低, 花粉常常出现 不育, 杂交结实较难。此外, 普通栽培种基因库狭 窄, 而具有丰富基因资源的野生种质难以与它们进 行有性杂交, 因而使外源基因的引进受到很大的限 制。近年来发展起来的体细胞融合技术为解决上述 问题开辟了一条新途径。在马铃薯体细胞融合的实 践中, Butenko 等(1980) [ 1 ]将栽培品种的原生质体 与 S. chacoense 的融合获得了抗马铃薯 Y 病毒的杂 种植株。此外, 马铃薯与二倍体野生种S. brev id ense ( 抗马铃薯卷叶病毒、Y 病毒和晚疫病) [ 4, 5 ]、S. p innatisectum (抗马铃薯晚疫病) [ 9 ]、S. bu lbocastanum (抗马铃薯晚疫病和线虫) [ 3, 6 ]和 S. circaeif olium (抗 马铃薯晚疫病) [ 8 ]等的原生质体融合也获得了杂种 植株。在获得双重经济性状的研究中, M attheij 等 (1992) [ 7 ]将马铃薯双单倍体品系与 S. p hu reja 的原 生质体融合, 从其杂种植株中选出了块茎产量比其 亲本高 3 倍的单株。这表明, 运用体细胞融合技术 能够获得有性杂交不亲和的种间体细胞杂种, 实现 有用性状向马铃薯栽培种的转移。我们通过近十年 的研究也表明体细胞融合是利用马铃薯野生种优良 基因的一条切实可行的途径[ 14 ]。 马铃薯种间体细胞杂种能否用于育种实践中, 取决于它们的育性、与栽培品种的可交配性和有益 性状的转移。我们所选用的三个融合亲本的育性都 比较高, 但由于它们之间亲缘关系较远, 有性杂交 2 期 司怀军等: 马铃薯种间体细胞杂种的育性和遗传改良 382 © 1995-2004 Tsinghua Tongfang Optical Disc Co., Ltd. All rights reserved